
 敬请关注公众号
敬请关注公众号

植物在生长过程中需要主动适应周围环境变化,该过程涉及到受体蛋白感受内外环境信号,而响应环境变化的受体是协调该过程的关键。细胞膜受体蛋白激酶FERONIA在植物中是个“多面手”,它不仅参与调控植物的有性生殖、细胞伸长、调节气孔开度和多种激素间的交叉会话,还调控植物响应外界逆境胁迫的应答过程。该特性使FERONIA成为近年来植物信号通路研究领域的“明星蛋白”。
位点信息
Locus: AT3G51550
Name: FERONIA
Other Name: FER
基因功能
CrRLK1L类受体激酶FER最早发现在植物双受精中发挥重要功能,随后研究人员发现FER参与调控各种生物活动,主要有:
(1) FER控制细胞伸长,fer-4 突变体的根毛、下胚轴、叶片、种皮等均表现出细胞伸长的缺陷;
(2) FER调控多种生物胁迫和非生物胁迫,fer-4 突变体对盐胁迫、渗透胁迫和温度胁迫均表现出敏感性。FER也参与植物的抗病反应;
(3) FER参与ABA、GA、BR、乙烯和生长素等激素响应,其中正调节BR和生长素信号,负调控ABA、乙烯等信号。
各种信号互相交叉,错综复杂,FER被认为是各种信号途径交叉会话的节点。
突变体信息
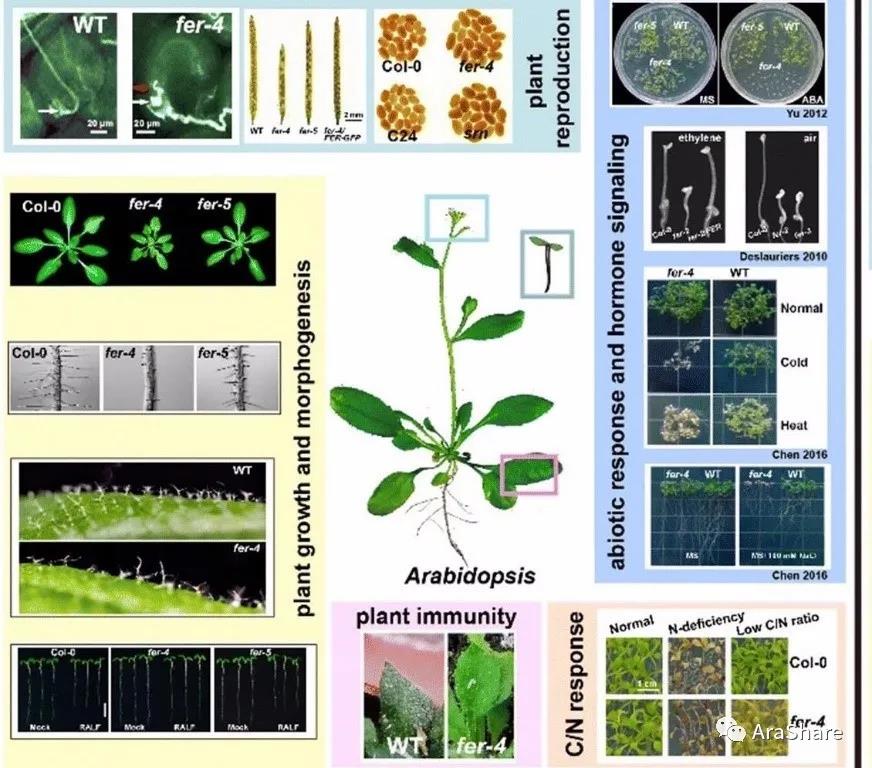
图1 fer突变体表型[1]
在国内拟南芥突变体共享中心AraShare中 (http://www.arashare.cn/) 有两个该基因的T-DNA插入突变体(N655026和N681032)。
基因表达谱
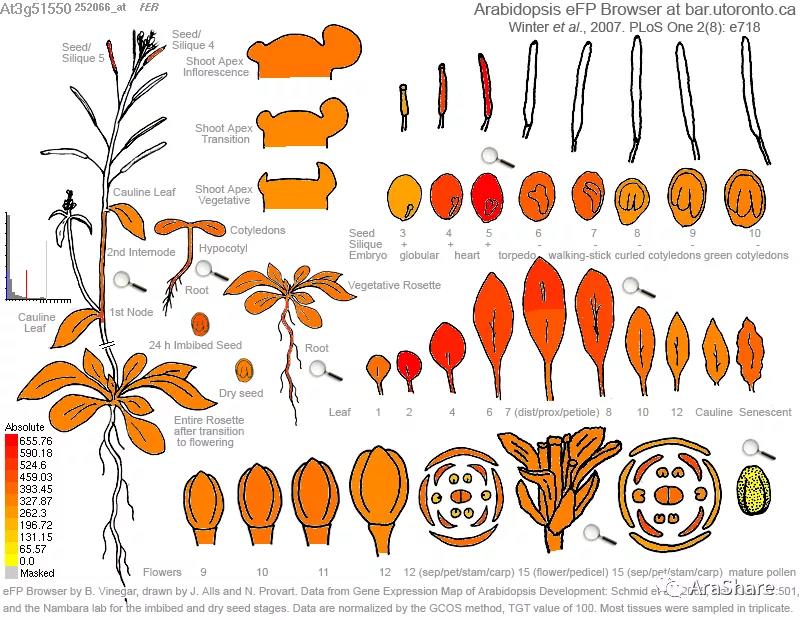
图2 FER基因表达图谱(引自eFP browser)
研究概况
2007年8月,UeliGrossniklaus课题组Juan-Miguel Escobar-Restrepo首次报道了fer 在生殖方面的表型,发表在Science杂志[2]。这就像一把钥匙,打开了FERONIA的金库。突变体fer 种荚结实率降低,但是雌雄配子体均没有发育缺陷。通过显微观察发现,该基因突变后,花粉管虽能准确到达珠孔,但不爆裂释放精细胞,在胚囊中不停止生长,最终授精失败引起不育。在西方神话寓意中,FERONIA代表了爱情女神。FERONIA基因也因其在植物生殖中的功能而得名。之后虽然每年均有多篇关于FER的文章报道,但是对于FER调控生殖发育的具体分子机理始终不太清楚。
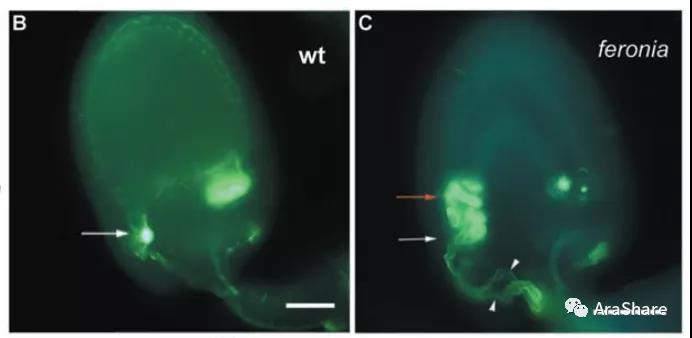
图3 FER参与花粉管破裂及释放精细胞[2]
直到2020年,美国马萨诸塞大学阿默斯特分校Alice Y Cheung和Henming Wu教授及山东农业大学的段巧红教授合作解析了fer突变体多个花粉管进入同一个胚珠的分子机理[3]。正常植物中,花粉管到达胚珠,便会破裂释放精细胞,而胚珠对于其他花粉管的吸引能力就会终止,最终只有一根花粉管进入珠孔。FERONIA调控了胚珠丝状器中低甲酯化果胶质的积累,进而调控了第一个花粉管诱导的一氧化氮NO在丝状器的积累。NO对诱导花粉管进入胚珠的诱饵蛋白进行亚硝基化修饰,阻止其分泌,也使其失去诱导其他花粉管的活性,从而保证其他花粉管不能进入这个“名花有主”的胚珠。而fer 突变体丝状器内的低甲酯化果胶质和NO明显降低,导致多个花粉管进入同一胚珠。FERONIA参与了胚珠协调“花粉管破裂”与“防止多个花粉管进入胚珠”两个重要的生物学过程,在植物受精中扮演着关键角色。
在此之前,段巧红教授已经发现并报道了FER在根毛极性生长中的作用机理。突变体fer-4 的根毛变短且形态异常,FER通过与分子开关小G蛋白ROP的活化因子GEF1互作,影响了ROP的活性及其下游活性氧ROS的产生,从而调控了根毛的极性生长过程[4]。FER不仅参与根毛的极性生长,还影响主根、下胚轴、叶片、种皮等多种组织的伸长。植物多肽激素RALF (快速碱化因子)能够结合FER并促进FER的磷酸化[5],磷酸化的FER招募RIPK激酶[6],启动FERONIA/RIPK之间的相互磷酸化,从而形成一个植物生长调节信号的“放大器”,最终影响 H+-ATPase的活性,抑制氢泵,导致细胞壁碱化抑制细胞伸长。
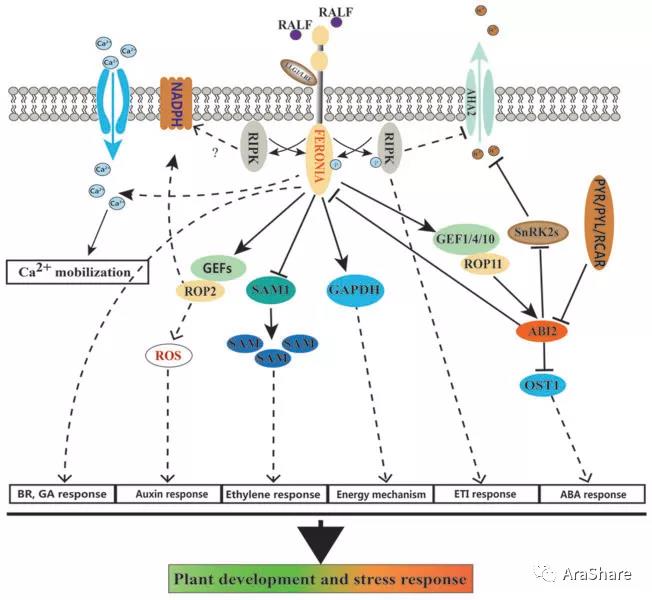
图4 FER参与植物生长和逆境胁迫[7]
FER还调控多种生物胁迫和非生物胁迫。2017年的science杂志报道,植物小肽RALF23等通过FER促进EFR/FLS2-BAK1复合体的生成,从而抑制植物的免疫反应[8]。2018年9月的Current Biology上报道了美国爱荷华州立大学尹延海教授发现的RALF23-FER-MYC2信号转导模块,FER通过磷酸化MYC2来调控其稳定性,抑制JA和COR信号转导,从而正向调节免疫[9]。此外,2018年PNAS上朱健康课题组的研究成果显示,细胞壁LRX蛋白通过与RALF多肽以及FER形成一个元件,感受盐胁迫造成的细胞壁变化,并将信号传递到胞内调控植物生长和耐盐性[10]。
近期,山东农业大学李厦教授课题组发现FER参与根的环形生长[11]。FER功能缺失增强了在倾斜平板上拟南芥根生长的波动性,在水平固体培养基表面进行反时针环形生长。利用NPA抑制生长素极性运输、引入PIN2或AUX1的突变体、抑制微管的动态组装和干扰钙离子进入,均能抑制这一环形生长,暗示FER通过生长素极性运输途径和影响微管动态组装及钙途径控制根的环形生长。
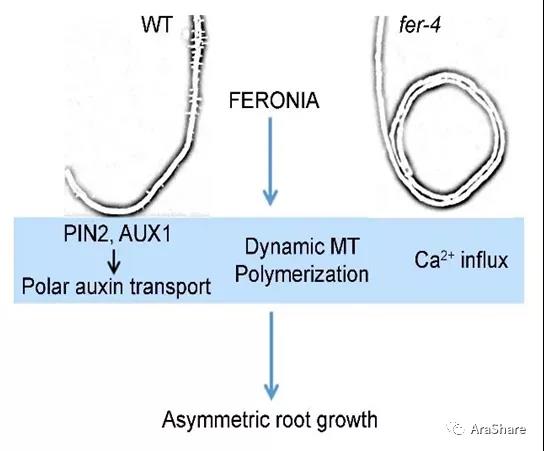
图5 FER抑制根回旋的工作模型[11]
基因网络
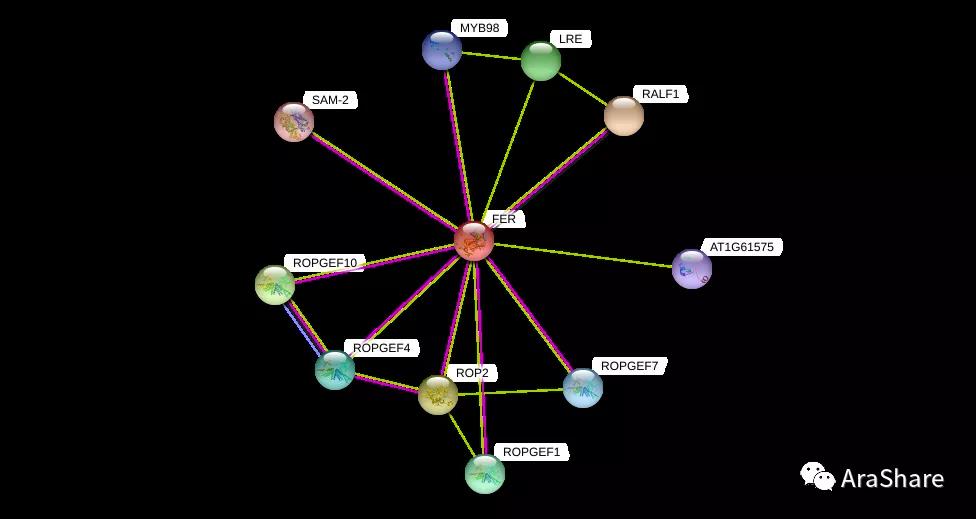
图6 拟南芥中FER蛋白互作网络预测(来自STRING)
其他物种研究情况简介
湖南大学于峰课题组鉴定了FER的水稻同源家族基因FLR,并获得多个FLR基因水稻突变体。研究发现flr突变体水稻稻瘟病抗性增强,但并不影响水稻的生长[12]。同时,该课题组揭示FLR通过与小G蛋白互作控制水稻种子的大小,解析了FLR在稻米品质中的作用[13]。另外,于峰课题组在根结线虫中鉴定了18种RALF类似编码基因,命名为RALF-like,线虫RALF-like能结合FER的胞外结构域,“挟持”植物中的FER信号途径,促进JA信号途径中核心蛋白MYC2的降解等过程,破坏植物的免疫系统[14]。FER在水稻中的同源蛋白也具有类似功能。这些工作为抗性作物育种设计奠定了基础。
展望
FERONIA被誉为植物界的“多面手”。一个蛋白能发挥如此多样性的功能,深得植物界的青睐。随着越来越多FER功能的发现,更多的科学问题也亟待解决。FER与RALF的识别机制是怎样的,哪些蛋白调控了RALF-FER的时空特异性表达,如何将现有的发现用于提高作物的环境适应性。相信随着生命科学的不断发展,科学家们最终会掀开女神FERONIA的神秘面纱。
参考文献
[1] Feng, Y. FERONIA cytoplasmic domain: node of varied signal outputs. aBIOTECH 1, doi:10.1007/s42994-020-00017-y (2020).
[2] Escobar-Restrepo, J. M. et al. The FERONIA receptor-like kinasemediates male-female interactions during pollen tube reception. Science 317, 656-660, doi:10.1126/science.1143562 (2007).
[3] Duan, Q. & Liu, M. J. FERONIAcontrols pectin- and nitric oxide-mediated male-female interaction. 579,561-566, doi:10.1038/s41586-020-2106-2 (2020).
[4] Duan, Q., Kita, D., Li, C., Cheung, A.Y. & Wu, H. M. FERONIA receptor-like kinase regulates RHO GTPase signalingof root hair development. Proceedings ofthe National Academy of Sciences of the United States of America 107, 17821-17826, doi:10.1073/pnas.1005366107 (2010).
[5] Haruta, M., Sabat, G., Stecker, K.,Minkoff, B. B. & Sussman, M. R. A peptide hormone and its receptor proteinkinase regulate plant cell expansion. Science 343, 408-411, doi:10.1126/science.1244454 (2014).
[6] Du, C. et al. Receptor kinase complex transmits RALF peptide signal toinhibit root growth in Arabidopsis. Proceedingsof the National Academy of Sciences of the United States of America 113, E8326-e8334, doi:10.1073/pnas.1609626113 (2016).
[7] Liao, H., Tang, R., Zhang, X., Luan,S. & Yu, F. FERONIA Receptor Kinase at the Crossroads of Hormone Signalingand Stress Responses.Plant & cellphysiology 58, 1143-1150, doi:10.1093/pcp/pcx048 (2017).
[8] Stegmann, M. et al. The receptor kinase FER is a RALF-regulated scaffoldcontrolling plant immune signaling. Science 355, 287-289, doi:10.1126/science.aal2541 (2017).
[9] Guo, H. et al. FERONIA Receptor Kinase Contributes to Plant Immunity bySuppressing Jasmonic Acid Signaling in Arabidopsis thaliana. Current biology : CB 28, 3316-3324.e3316, doi:10.1016/j.cub.2018.07.078 (2018).
[10] Zhao, C. et al. Leucine-rich repeat extensin proteins regulate plant salttolerance in Arabidopsis. 115, 13123-13128, doi:10.1073/pnas.1816991115 (2018).
[11] Li, E., Wang, G., Zhang, Y. L., Kong, Z.& Li, S. FERONIA mediates root nutating growth. doi:10.1111/tpj.14984 (2020).
[12] Yang, Z. et al. Mutations of two FERONIA-like receptor genes enhance riceblast resistance without growth penalty. Journalof experimental botany 71,2112-2126, doi:10.1093/jxb/erz541 (2020).
[13] Wang, L. et al. Roles of FERONIA-like receptor genes in regulating grainsize and quality in rice. Science China.Life sciences, doi:10.1007/s11427-020-1780-x (2020).
[14] Zhang, X. et al. Nematode-Encoded RALF Peptide Mimics Facilitate Parasitismof Plants through the FERONIA Receptor Kinase. Molecular plant13,1434-1454, doi:10.1016/j.molp.2020.08.014 (2020).
本推文基于科学文献和网络资讯写作而成,未经领域内专家审阅;限于笔者学识水平,难免存在错漏,仅供参考!