
 敬请关注公众号
敬请关注公众号

SPL(SQUAMOSA PROMOTER BINDING PROTEIN-LIKE)作为转录因子,参与到植物生长发育的各个阶段,从根的生长、花发育及果实成熟,覆盖面非常广泛。而作为SPL家族的中间儿,关于SPL8的研究却不是很多,大多集中在其参与到花器官发育的调控过程。但是SPL8并不丰富的“人生”经历同样也可以给我们带来一些启示。
基因信息
Locus:AT1G02065
Name: SPL8
基因功能
SPL8基因可以参与发育和花器官发生的,包括胚珠分化、花粉产生、花丝伸长、种子形成和角果伸长;似乎也在萼片毛状体的形成中起作用;同时也可能对花中赤霉素(GA)信号有正向调节作用。
突变体信息
SPL8突变体会影响大小孢子的发生,导致萼片上毛状态的形成以及使雄蕊丝生长方面受到影响。在国内拟南芥突变体共享中心AraShare中有一个该基因的T-DNA插入突变体(N692410,详情请查阅http://www.arashare.cn/)。

图1 spl8突变体与野生型表型
基因表达谱
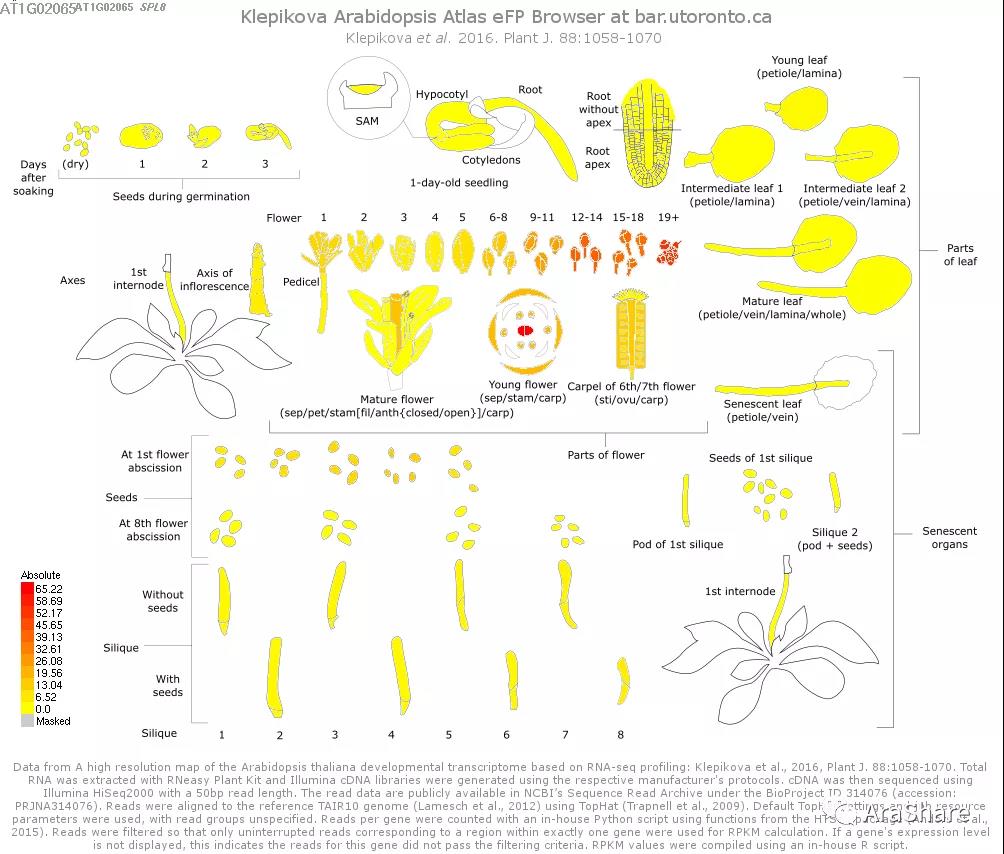
图2 拟南芥SPL8基因表达谱
研究概况
SPL家族是个庞大的家族,在拟南芥中共鉴定到16个成员,SPL8作为该家族中的中间成员,并没有像其他兄弟姐妹一样受到太多的关注,大多数时候都是在研究其他成员时顺带上它,为它量身定做的研究并不多,因而SPL8一直四处游荡,居无定所。直到2003年德国马克思普朗克植物育种研究所在the Plant Cell发表研究论文“SPL8, an SBP-Box Gene That Affects Pollen Sac Development in Arabidopsis”才有了SPL8自己的“房子”。在该研究论文中,Ulrike S. Unte等人利用拟南芥的转座子诱变种群来寻找SBP-box基因的插入,在SPL8基因中鉴定和分离了三个这样的转座子插入等位基因。相应的突变植株(spl8-1,spl8-2,spl8-3)都表现出很强的生育力下降,主要是由于发育中的花药细胞分化异常的结果。然而,SPL8的缺失并不会在很大程度上影响拟南芥的生长,因为所有的spl突变体都可以产生有活性的花粉和种子,这可能是因为发生了功能冗余的情况,也正因此,SPL8才一直被遗忘。但是对SPL家族进行蛋白序列比对发现,其他成员除了SBP-box区域和SPL8高度同源以外,其他部分并无太多相似性,因此对SBP-box区域的功能研究依旧是个值得探索的方向[1]。
SPL8的房子搭建好以后,同样是来自德国马克思普朗克植物育种研究所的研究人员开始了“装修”事宜。首先在2006年,Yan Zhang等人带来了赤霉素(GA),一种植物生长调节剂,调节植物的许多发育过程,包括种子萌发、根和茎的伸长、莲座叶扩张、花的诱导和花药的发育。通过SPL8的突变实验及多种生化手段得到SPL8的缺失会导致GA的不敏感性的结果,同时也通过SPL8功能获得实验反向证明了该结果[2]。时间来到2013年,Shuping Xing等人则把BIM1带到了SPL8身旁,并通过原位杂交、功能获得等试验证明了BIM1可以增强SPL8半不育的表型,同时也得出BIM1和SPL8一样会影响花药早期的发育的结果[3]。定睛一看,给SPL8建房子和进行装修的都是Peter Huijser带来的人手。感谢他们为SPL8所做的一切。

图3 sp8和bim1 对植株生长的影响
其他物种研究情况简介
2019年王增裕团队在New Phytologist在线发表了研究论文“SPL7 and SPL8 represent a novelflowering regulation mechanism in switchgrass”,揭示了过表达SPL7和SPL8会促进柳枝稷开花,而下调单个基因则在一定程度上延迟了开花。同时发现SPL8通过直接上调SEP3和MADS32基因诱导开花[4]。在此之前,同样是王增裕团队发现在苜蓿中SPL8的下调会增加其抗胁迫水平,同时SPL8还可以影响腋芽和分枝的发育[5]。除此之外,2017年来自山西农业大学的张晶等人对白菜(Brassica rapa subsp. chinensis)中的SPL8基因进行了鉴定,并得出BrcSPL8在白菜花发育过程中发挥作用,有助于白菜花发育机制的研究的结论[6]。
展望
目前对SPL8的研究的确不够深入,乍一看是劣势,仔细思索一番,却发现这也正是它的优势所在。因为SPL家族中其他成员研究的深入,为SPL8的研究提供了扎实而又丰富的基础,尤其是在SBP-box区域功能方面。同时无论是在草本中还是林木中,SPL8的研究空间也很广阔。相信在不久的将来,会陆续有关于SPL8的研究报道出来,SPL8的“房子”会越来越漂亮。
参考文献
[1] U.S. Unte, A. Sorensen, P.Pesaresi, M. Gandikota, D. Leister, H. Saedler, P. Huijser, SPL8, anSBP-Box Gene That Affects Pollen Sac Development in Arabidopsis, The PlantCell, 15 (2003) 1009-1019.
[2] Y. Zhang, S. Schwarz, H.Saedler, P. Huijser, SPL8, a local regulator in a subset ofgibberellin-mediated developmental processes in Arabidopsis, PLANT MOL BIOL,63 (2006) 429-439.
[3] S. Xing, V. Quodt, J. Chandler,S. Hohmann, R. Berndtgen, P. Huijser, SPL8 Acts Together with theBrassinosteroid-Signaling Component BIM1 in Controlling Arabidopsisthaliana Male Fertility, Plants (Basel), 2 (2013) 416-28.
[4] J.Gou, C. Tang, N. Chen, H. Wang, S. Debnath, L. Sun, A. Flanagan, Y. Tang, Q.Jiang, R.D. Allen, Z.Y. Wang, SPL7 and SPL8 represent a novelflowering regulation mechanism in switchgrass, NEW PHYTOL, 222 (2019)1610-1623.
[5] J. Gou, S. Debnath, L. Sun, A.Flanagan, Y. Tang, Q. Jiang, J. Wen, Z. Wang, From model to crop: functionalcharacterization of SPL8 in M. truncatula led to genetic improvement ofbiomass yield and abiotic stress tolerance in alfalfa, PLANT BIOTECHNOL J,16 (2018) 951-962.
[6] J. Zhang, A. Ping, X. Wang, G.Li, Z. Zhu, M. Li, G. Xing, L. Hou, Cloning and expression analysis of SPL8homolog from pak choi ( Brassica rapa subsp. chinensis ), Biotechnology,biotechnological equipment, 31 (2017) 1132-1138.
本推文基于科学文献和网络资讯写作而成,未经领域内专家审阅;限于笔者学识水平,难免存在错漏,仅供参考!