
 敬请关注公众号
敬请关注公众号

Receptor for Activated C Kinase 1 (RACK1) 是一类色氨酸--天门冬氨酸(WD40)重复结构蛋白。在多种哺乳动物生长发育过程关键调控作用。拟南芥中该蛋白家族含有3个编码RACK1的基因,氨基酸水平相似性相较哺乳动物RACK1超过75%,植物的RACK1蛋白在信号转导、免疫等多个方面都扮演重要角色[1]。
位点信息
1. AT1G18080; Name: ATARCA, ATRACK1, RACK1A, RACK1A_AT, RECEPTOR FOR ACTIVATED C KINASE 1 A, SAC53, SUPPRESSOR OF ACAULIS 53
2. AT1G48630; Name: RACK1B, RACK1B_AT, RECEPTOR FOR ACTIVATED C KINASE 1B
3. AT3G18130; Name: RACK1C, RACK1C_AT, RECEPTOR FOR ACTIVATED C KINASE 1C
基因功能
RACK1编码一个WD40重复蛋白结构的脚手架蛋白。在细胞核和内质网定位,由于其受到多种蛋白修饰,与多种信号组分重要蛋白相互作用,囊括细胞壁生物合成、 光合作用、细胞骨架、免疫、非生物胁迫、信号传导、转录水平调控、核糖体生物合成、蛋白质合成、生长发育调控及激素响应,未来随着对RACK1蛋白的研究可能拓展该蛋白更多的生物学功能。
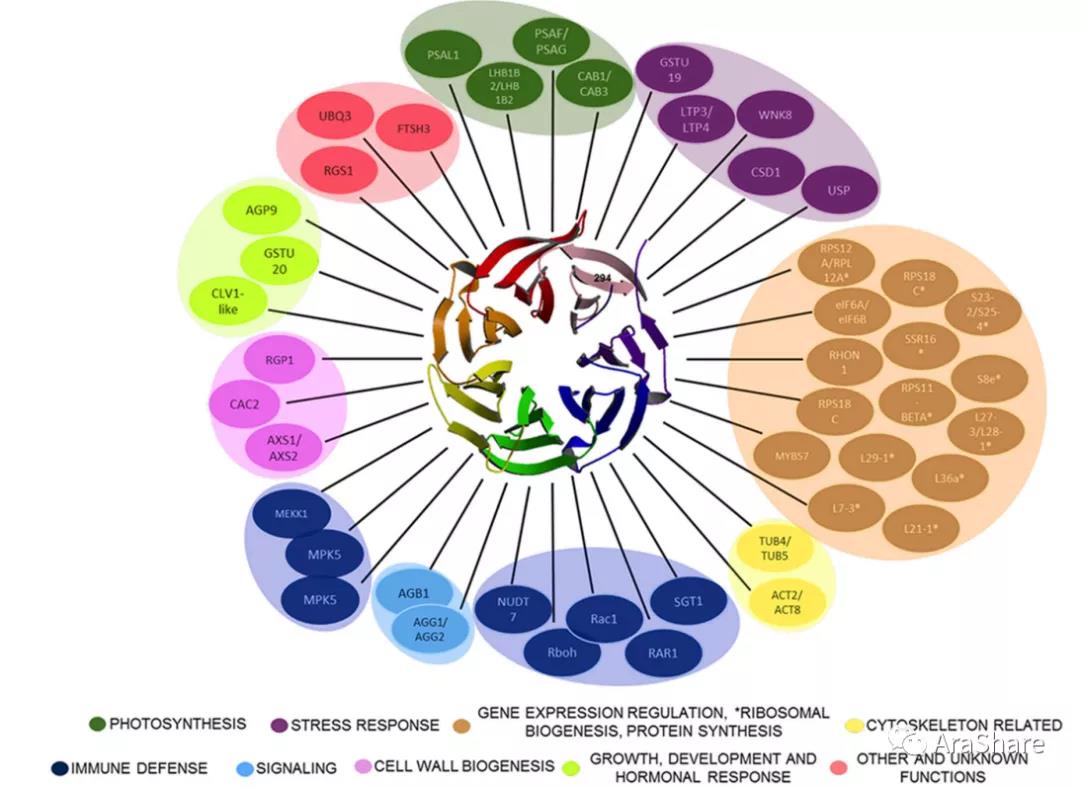
图1 RACK1通过与其互作配体参与多种细胞调控过程[2]
基因表达谱
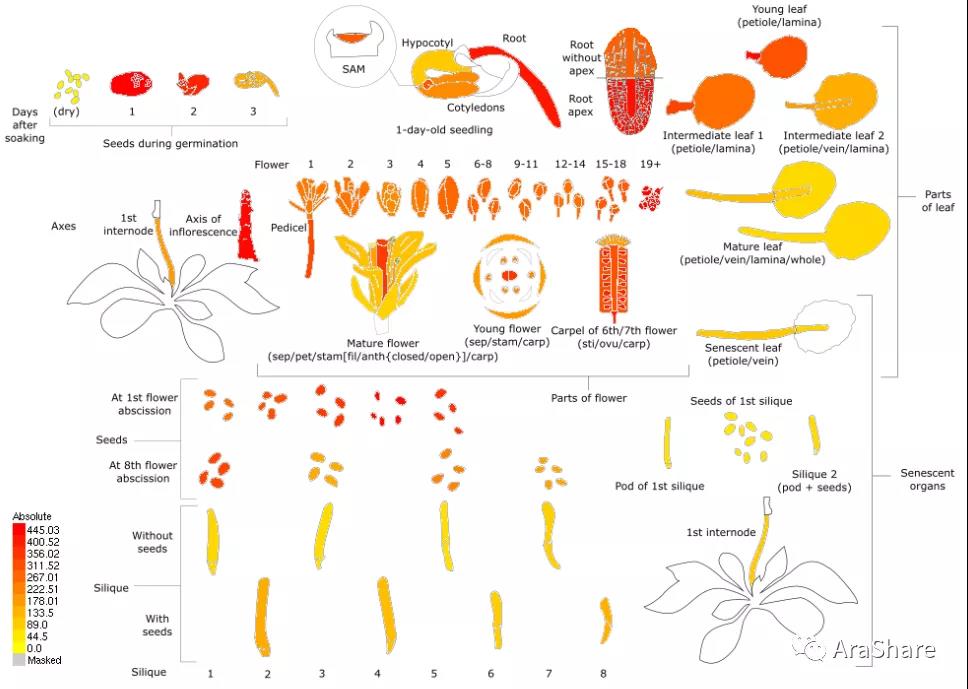
图2 拟南芥中RACK1A基因表达谱(Source: eFP Broswer from http://www.bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi)
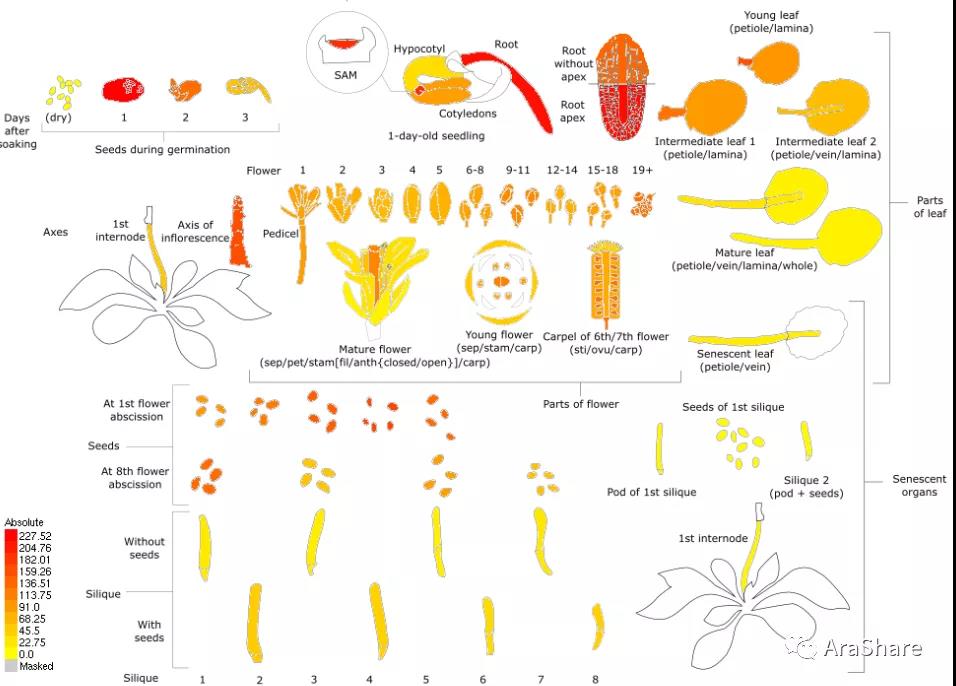
图3 拟南芥中RACK1B基因表达谱(Source: eFP Broswer from http://www.bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi)
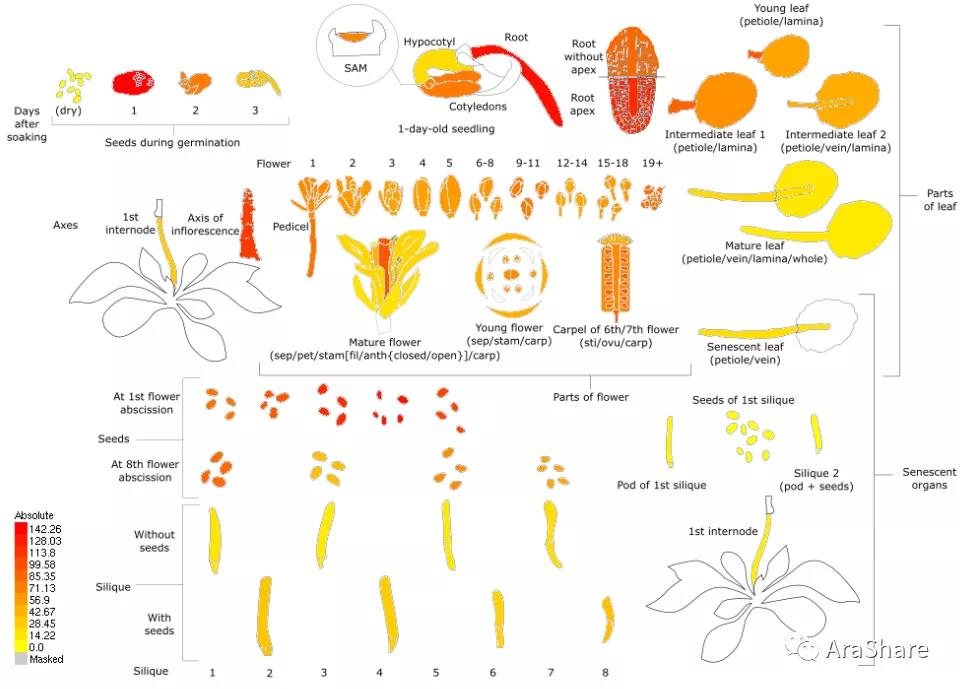
图4 拟南芥中RACK1C基因表达谱(Source: eFP Broswer from http://www.bar.utoronto.ca/efp/cgi-bin/efpWeb.cgi)
从图上我们可以看出,三个不同的RACK1同源蛋白在拟南芥的转录水平依次是RACK1A>RACK1B>RACK1C,功能上这三个蛋白的重要性也是按照这样排序。
研究概况
RACK1蛋白首先从动物被发现,第一个植物RACK1基因从烟草By-2细胞系中克隆,命名为arcA[3]。随后在其他物种都有相关报道。由于动物中RACK1蛋白只有一种,而植物则有不止一种,目前认为植物RACK1蛋白可能存在冗余。
从结构上看,拟南芥RACK1与G蛋白β相似,存在7片类似螺旋桨结构。每个螺旋桨围绕一个假想轴排列,并且中央存在一个直径为9 Å水分子通道[1]。作为支架蛋白,RACK1A存在2个保守的蛋白质相互作用位点,这两个位点分别位于RACK1螺旋桨顶部和底部,说明其可能通过顶对顶(top-to-top)顶对底(top-to-bottom)或者底对底(bottom-to-bottom)发生相互作用。
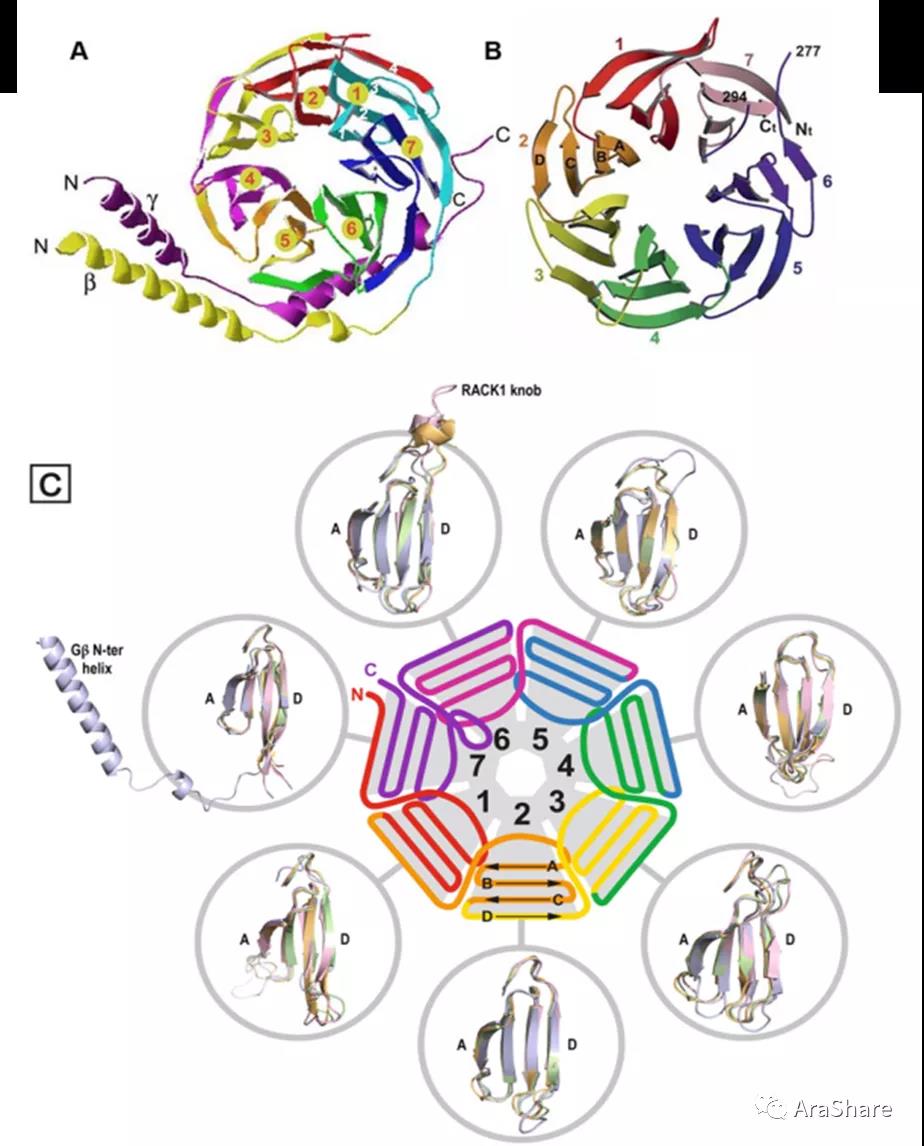
图5 拟南芥Gβγ和RACK1的结构。(A) Gβγ; (B) RACK1; (C) RACK1蛋白7个螺旋桨结构[1,4,5]
由于在动物研究中,RACK1蛋白在GPCR途径调控过程研究非常深入,受到大量关注。后续研究发现其在核糖体结构上交联的证据。拟南芥RACK1A蛋白可以与40S小亚基结合。RACK1与活化形式的蛋白激酶C(PKC)结合, 将PKC定位到40S核糖体上并磷酸化翻译起始因子eIF6[6]。2014年,德国图宾根大学的Sascha Laubinger课题组在Plant Journal报道了题为“RACK1 scaffold proteins influence miRNA abundance in Arabidopsis“的研究论文。他们发现在rack1突变体中miRNA水平降低,并且生化证据表明RACK1与miRNA加工AGO1复合体的SE蛋白互作,证明rack1突变体会造成miRNA的靶mRNA过量积累[7]。
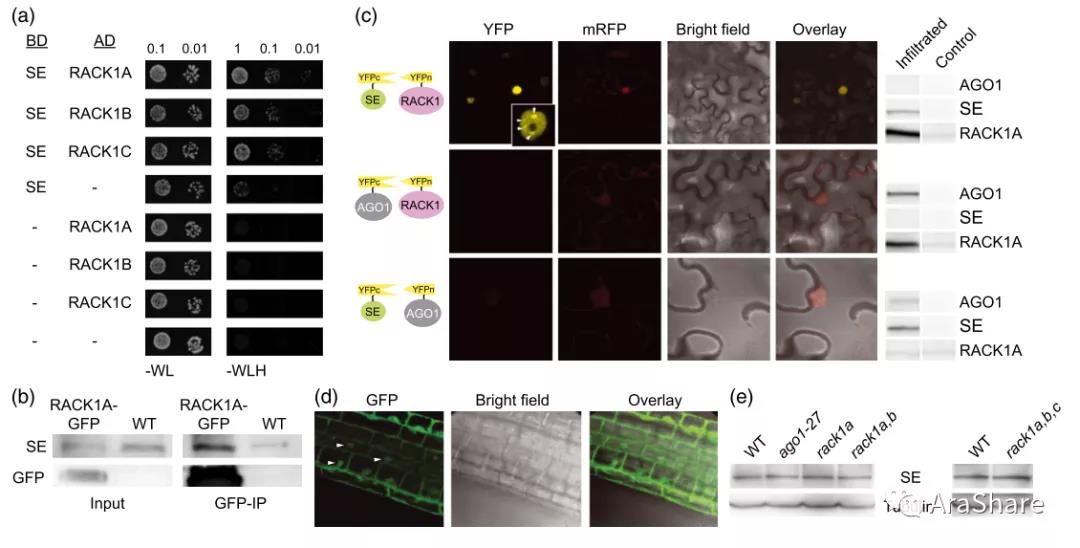
图6 RACK1蛋白与SE蛋白互作证据[7]
2015年,陈金桂博士和Alan M. Jones课题组在Plant Physiology发表题为“ Arabidopsis Receptor of Activated C Kinase1 Phosphorylation by WITH NO LYSINE8 KINASE”的研究论文,为RACK1A的蛋白修饰提供了重要的生化证据。
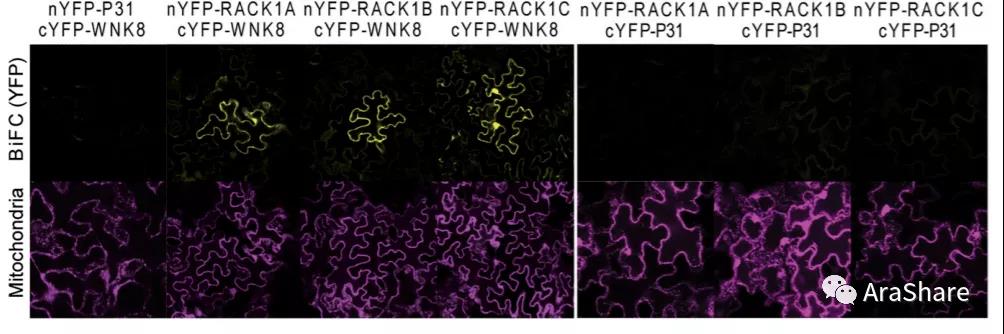
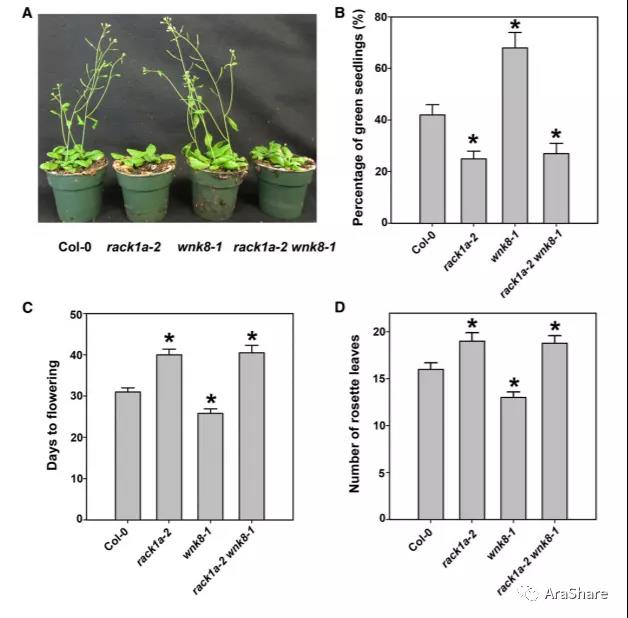
图7 RACK1与WNK8互作生化证据和遗传证据[8]
真正把RACK1跟抗病信号调控通路联系起来的是2015年Frederick M. Ausubel课题组发表在Nature上的题为“Pathogen-secreted proteases activate a novel plant immune pathway”的研究论文。MAPK信号通路级联放大是动植物中普遍存在,而在拟南芥,信号感知是如何传导到MAPK激活下游植物免疫的分子机制仍不清楚。这篇文章发现病原体分泌蛋白可以激活由RACK1支架蛋白介导的涉及Gα,Gβ和Gγ异源复合体这些蛋白相互作用激活了病原体分泌蛋白酶-G蛋白-RACK1-MAPK级联模块信号,并且这个调控信号与菌鞭毛蛋白引起的免疫信号通路又不尽相同。这篇文章用扎实的把RACK1在植物免疫调控过程的重要作用揭示出来[9]。美国德州农工大学的何平教授也在Molecular Plant 发表评论“Stack Heterotrimeric G Proteins and MAPK Cascades on a RACK”[10]。浙江大学张舒群教授也在Trends in Plant Science发表题为“RACK1, scaffolding a heterotrimeric G protein and a MAPK cascade”评论[11]。
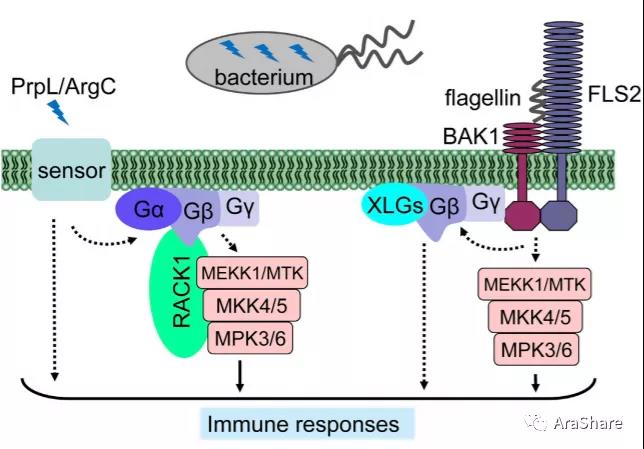
图8 异源G蛋白复合体信号传导参与植物先天免疫调控过程[10]
2019年,东北师范大学王树才课题组(今临沂大学)在Plant Science发表题为“Integration of RACK1 and ethylene signaling regulates plant growth and development in Arabidopsis”研究论文,揭示了虽然RACK1不与乙烯信号途径关键调控因子Ethylene INsensitive 2(EIN2)发生相互作用,但是遗传上和表型都有直接关联[12]。同年在Plant Signaling & Behavior杂志报道了RACK1蛋白可以受到盐信号和ABA信号,通过调控miR393s的发生抑制生长素受体TIR1/AFB2信号调控下游生长发育过程[13]。
上述研究论文多数关于RACK1A蛋白,而关于RACK1B蛋白,2017年,孙卫宁课题组在Scientific Reports报道了拟南芥ABA信号响应过程,RACK1B受到SUMO化修饰,并提高了与RAP2.6蛋白互作的稳定性[14]。
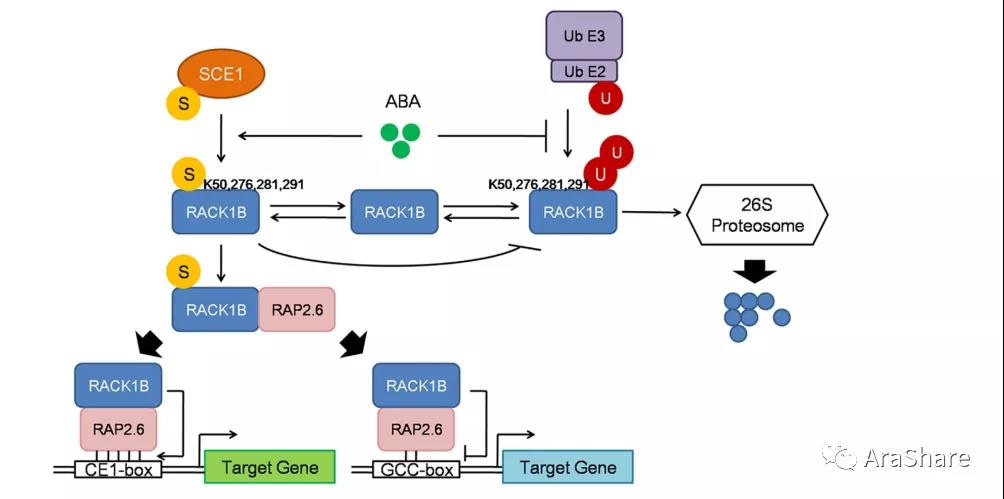
图9 ABA信号通过SUMO化RACK1B增加其与RAP2.6蛋白稳定性示意图[14]
突变体信息
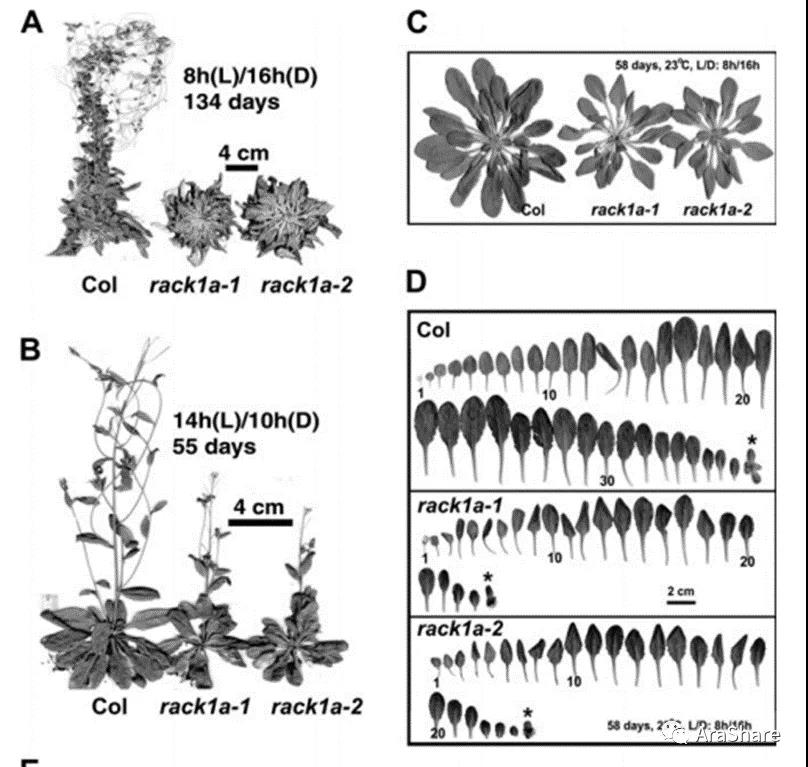
图10 rack1a-1,rack1a-2突变体植株及叶片表型[15]

图11 rack1各拟南芥突变体ABA处理表型图[16]

图12 多种rack1突变体杂交组合表型[17]
rack1a突变体在Alan M. Jones课题组于2006年在Journal of Experimental Botany发表的题为“RACK1 mediates multiple hormone responsiveness and developmental processes in Arabidopsis”研究论文展现了其多种表型,相较于野生型Col-0,rack1a突变体表现出包括子叶发育不良、莲座叶数量增加、叶片发育畸形、发育-生殖周期转换推迟等表型[15]。本文第一作者陈金桂博士于2009年在Journal of Experimental Botany发表的题为“ RACK1 is a negative regulator of ABA responses in Arabidopsis”也展现多个rack1突变体ABA处理和发芽等表型[16]。2010年,陈金桂博士继续在BMC Plant Biology报道了rack1的突变体表型。他的结果表明RACK1A/1B/1C存在功能不平等的冗余,三者表达模式相似,而且还能互相调节彼此表达[17]。
目前rack1a突变体在国内拟南芥突变体中心(AraShare)有一份材料SN23374(SALK_099406C),rack1b突变体在有一份材料L0899(SAIL_413_B07),rack1c突变体有一份材料SNKF4070 (SALK_017913C)。
其他物种研究进展
近年来,对于RACK1蛋白在其他植物物种的报道包括棉花[18]、本式烟草[19]、大豆[20]、羽衣甘蓝[21]、玉米[22]、水稻[23–25]。
早在2008年,日本奈良先端科学技术大学院大学的Ko Shimamoto课题组在Plant Cell报道了水稻OsRACK1A蛋白与Rac1免疫复合体互作,揭示了RACK1调控水稻先天性免疫途径在植物界可能都是保守的[17]。而2010年后,水稻的这三篇文章分别从ABA、耐盐、与CONSTAN-like9蛋白互作调控SA和乙烯途径等角度多方面解析OsRACK1的蛋白功能。
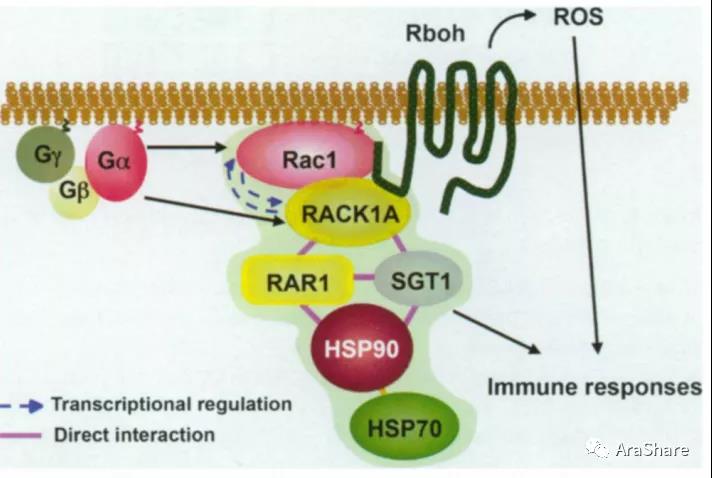
图13 RACK1A介导的水稻先天免疫调控过程示意图[17]
2019年,中科院上海卓越创新中心林鸿宣课题组在Molecular Plant发表了题为“Translational Regulation of Plant Response to High Temperature by a Dual-Function tRNAHis Guanylyltransferase in Rice”的研究论文。这篇研究论文发现AET1-eIF3h-RACK1三者相互作用,共同调控含有uORF结构的ARF基因蛋白翻译过程。在本文作者认为水稻RACK1蛋白在这起了招募这些蛋白的作用,这三者(或者有更多其他核糖体蛋白)共同形成复合体发挥核糖体调控蛋白翻译过程[26]。关于RACK1蛋白近年来陆续报道一些植物综述,读者有兴趣可以关注[27–31]。
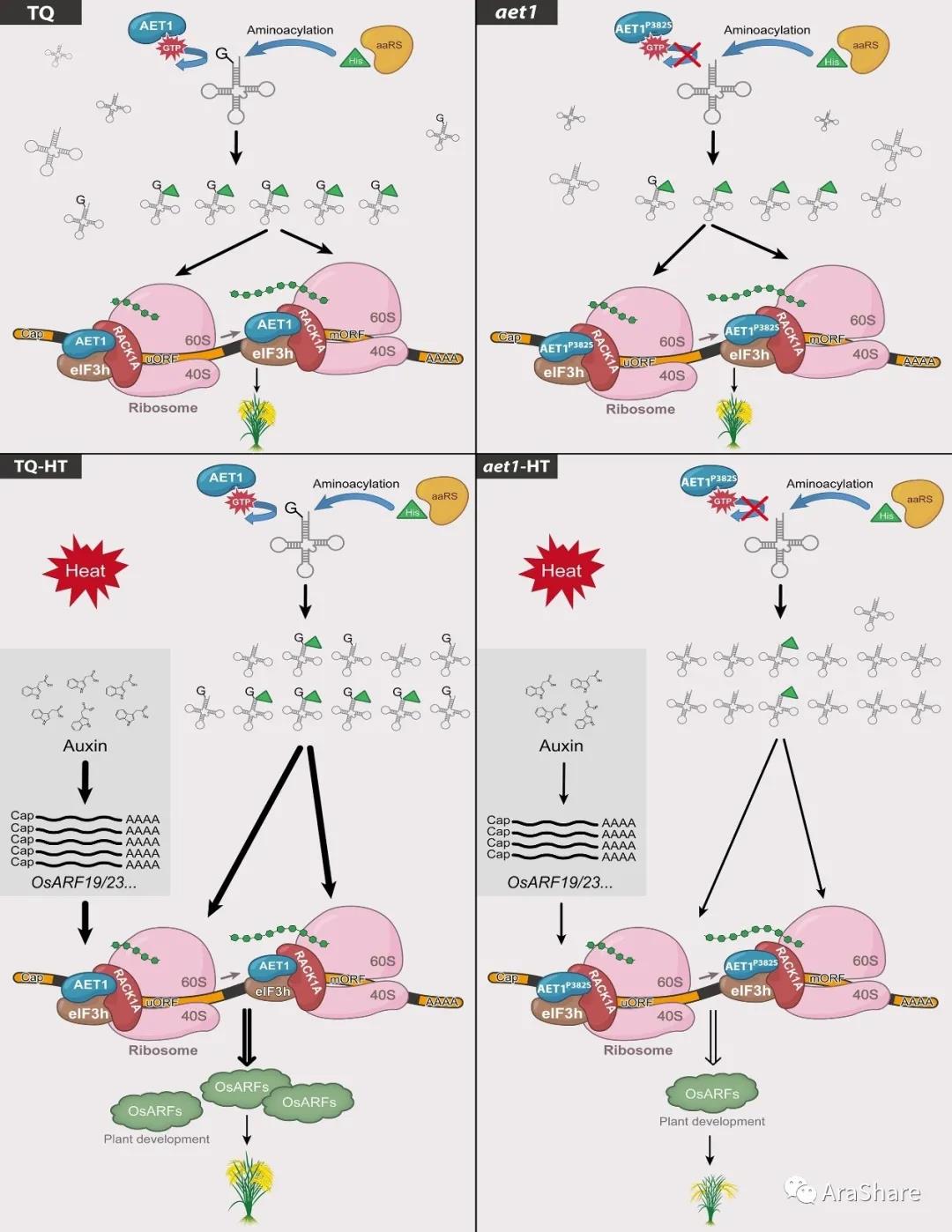
图14 AET1-RACK1A-eIF3h三者互作共同调控水稻高温适应性分子机制示意图[26]
展望
对于像RACK1这一类WD40支架蛋白,其蛋白具有功能多样性,协调和整合多种信号通路发挥重要作用。未来笔者觉得对于这类基因除了继续挖掘其相互作用的对象寻找不同的重要信号通路参与者,挖掘他们的调控时空特异性外,还可以挖掘上游对于RACK1的转录因子调控、下游对RACK1蛋白多种修饰(磷酸化等)。逐步找出RACK1蛋白表型与某些特殊调控的表型和生化关联。并最终能够寻求其在免疫、生长发育等过程的应用。
参考文献
[1] Dahong, L.; Dongping, Z.; Dandan, C.; Jiansheng, L. Research Advances in Plant RACK1 Proteins. CHINESE Bull. Bot. 2011, 46, 224–232.
[2] Islas-Flores, T.; Rahman, A.; Ullah, H.; Villanueva, M.A. The receptor for activated C kinase in plant signaling: Tale of a promiscuous little molecule. Front. Plant Sci. 2015, 6, 1–19.
[3] Ishida, S.; Takahashi, Y.; Nagata, T. Isolation of cDNA of an auxin-regulated gene encoding a G protein β subunit-like protein from tobacco BY-2 cells. Proc. Natl. Acad. Sci. U. S. A. 1993, 90, 11152–11156.
[4] Ullah, H.; Scappini, E.L.; Moon, A.F.; Williams, L.V.; Armstrong, D.L.E.E. Structure of a signal transduction regulator, RACK1, from Arabidopsis thaliana. Protein Sci. 2008, 17, 1771–1780.
[5] Chen, S.; Spiegelberg, B.D.; Lin, F.; Dell, E.J.; Hamm, H.E. Interaction of Gβγ with RACK1 and other WD40 repeat proteins. J. Mol. Cell. Cardiol. 2004, 37, 399–406.
[6] Chang, I.F.; Szick-Miranda, K.; Pan, S.; Bailey-Serres, J. Proteomic characterization of evolutionarily conserved and variable proteins of arabidopsis cytosolic ribosomes. Plant Physiol. 2005, 137, 848–862.
[7] Speth, C.; Willing, E.M.; Rausch, S.; Schneeberger, K.; Laubinger, S. RACK1 scaffold proteins influence miRNA abundance in Arabidopsis. Plant J. 2013, 76, 433–445.
[8] Urano, D.; Czarnecki, O.; Wang, X.; Jones, A.M.; Chen, J.G. Arabidopsis receptor of activated c kinase1 phosphorylation by WITH NO LYSINE8 KINASE. Plant Physiol. 2015, 167, 507–516.
[9] Cheng, Z.; Li, J.F.; Niu, Y.; Zhang, X.C.; Woody, O.Z.; Xiong, Y.; Djonović, S.; Millet, Y.; Bush, J.; McConkey, B.J.; et al. Pathogen-secreted proteases activate a novel plant immune pathway. Nature 2015, 521, 213–216.
[10] Meng, X.; Shan, L.; He, P. Stack Heterotrimeric G Proteins and MAPK Cascades on a RACK. Mol. Plant 2015, 8, 1691–1693.
[11] Su, J.; Xu, J.; Zhang, S. RACK1, scaffolding a heterotrimeric G protein and a MAPK cascade. Trends Plant Sci. 2015, 20, 405–407.
[12] Wang, W.; Wang, X.; Wang, X.; Ahmed, S.; Hussain, S.; Zhang, N.; Ma, Y.; Wang, S. Integration of RACK1 and ethylene signaling regulates plant growth and development in Arabidopsis. Plant Sci. 2019, 280, 31–40.
[13] Denver, J.B.; Ullah, H. miR393s regulate salt stress response pathway in Arabidopsis thaliana through scaffold protein RACK1A mediated ABA signaling pathways. Plant Signal. Behav. 2019, 14.
[14] Guo, R.; Sun, W. Sumoylation stabilizes RACK1B and enhance its interaction with RAP2.6 in the abscisic acid response. Sci. Rep. 2017, 7, 1–13.
[15] Chen, J.G.; Ullah, H.; Temple, B.; Liang, J.; Guo, J.; Alonso, J.M.; Ecker, J.R.; Jones, A.M. RACK1 mediates multiple hormone responsiveness and developmental processes in Arabidopsis. J. Exp. Bot. 2006, 57, 2697–2708.
[16] Guo, J.; Wang, J.; Xi, L.; Huang, W.D.; Liang, J.; Chen, J.G. RACK1 is a negative regulator of ABA responses in arabidopsis. J. Exp. Bot. 2009, 60, 3819–3833.
[17] Nakashima, A.; Chen, L.; Nguyen, P.T.; Fujiwara, M.; Hann, L.W.; Kuwano, M.; Umemura, K.; Shirasu, K.; Kawasaki, T.; Shimamotoa, K. RACK1 functions in rice innate immunity by interacting with the Rac1 immune complex. Plant Cell 2008, 20, 2265–2279.
[18] Shu, F.; Wang, B.; Liu, D.; Wang, X.; Zhang, B. Functional characterization of a receptor for activated C kinase 1 (RACK1) gene from upland cotton (Gossypium hirsutum L.). Plant Growth Regul. 2020, 91, 359–369.
[19] Hyodo, K.; Suzuki, N.; Okuno, T. Hijacking a host scaffold protein, RACK1, for replication of a plant RNA virus. New Phytol. 2019, 221, 935–945.
[20] Li, D.H.; Chen, F.J.; Li, H.Y.; Li, W.; Guo, J.J. The Soybean GmRACK1 Gene Plays a Role in Drought Tolerance at Vegetative Stages. Russ. J. Plant Physiol. 2018, 65, 541–552.
[21] Li, D.H.; Shen, F.J.; Li, H. yan; Li, W. Kale BoRACK1 is involved in the plant response to salt stress and Peronospora brassicae Gaumann. J. Plant Physiol. 2017, 213, 188–198.
[22] Wang, B.; Yu, J.; Zhu, D.; Chang, Y.; Zhao, Q. Maize ZmRACK1 is involved in the plant response to fungal phytopathogens. Int. J. Mol. Sci. 2014, 15, 9343–9359.
[23] Zhang, D.; Chen, L.; Li, D.; Lv, B.; Chen, Y.; Chen, J.; Yan, X.; Liang, J. OsRACK1 is involved in abscisic acid- and H2O2- mediated signaling to regulate seed germination in rice (Oryza sativa, L.). PLoS One 2014, 9, 1–11.
[24] Liu, H.; Dong, S.; Sun, D.; Liu, W.; Gu, F.; Liu, Y.; Guo, T.; Wang, H.; Wang, J.; Chen, Z. CONSTANS-like 9 (OsCOL9) interacts with receptor for activated C-kinase 1(OsRACK1) to regulate blast resistance through salicylic acid and ethylene signaling pathways. PLoS One 2016, 11, 1–17.
[25] Zhang, D.; Wang, Y.; Shen, J.; Yin, J.; Li, D.; Gao, Y.; Xu, W.; Liang, J. OsRACK1A, encodes a circadian clock-regulated WD40 protein, negatively affect salt tolerance in rice. Rice 2018, 11, 1–15.
[26] Chen, K.; Guo, T.; Li, X.M.; Zhang, Y.M.; Yang, Y.B.; Ye, W.W.; Dong, N.Q.; Shi, C.L.; Kan, Y.; Xiang, Y.H.; et al. Translational Regulation of Plant Response to High Temperature by a Dual-Function tRNAHis Guanylyltransferase in Rice. Mol. Plant 2019, 12, 1123–1142.
[27] Miller, J.C.; Chezem, W.R.; Clay, N.K. Ternary WD40 repeat-containing protein complexes: Evolution, composition and roles in plant immunity. Front. Plant Sci. 2016, 6.
[28] Islas-Flores, T.; Rahman, A.; Ullah, H.; Villanueva, M.A. The receptor for activated C kinase in plant signaling: Tale of a promiscuous little molecule. Front. Plant Sci. 2015, 6.
[29] Adams, D.R.; Ron, D.; Kiely, P.A. RACK1, A multifaceted scaffolding protein: Structure and function. Cell Commun. Signal. 2011, 9.
[30] Nielsen, M.H.; Flygaard, R.K.; Jenner, L.B. Structural analysis of ribosomal RACK1 and its role in translational control. Cell. Signal. 2017, 35, 272–281.
[31] Racchi, M.; Buoso, E.; Ronfani, M.; Serafini, M.M.; Galasso, M.; Lanni, C.; Corsini, E. Role of hormones in the regulation of RACK1 expression as a signaling checkpoint in immunosenescence. Int. J. Mol. Sci. 2017, 18, 1–12.
本推文基于科学文献和网络资讯写作而成,未经领域内专家审阅;限于笔者学识水平,难免存在错漏,仅供参考!