
 敬请关注公众号
敬请关注公众号

与动物不同,植物在环境中的位置是相对固定的。在复杂的环境中,植物面临多种病原物的威胁,包括细菌,真菌,卵菌,病毒和线虫等。在农业生产中,每年都因病害而导致大量的经济损失。面对这些威胁,植物进化出了一套复杂的防卫网络,而NPR1是该网络中最重要的调控基因之一。
位点信息
Locus:AT1G64280
Name:NPR1(NONEXPRESSER OF PR GENES 1)
Other Name:NIM1,SAI1
基因功能
植物的免疫反应与生长发育往往是拮抗的,持续的抗病反应无端地消耗太多能量,而过头的抗病反应则直接导致植物的衰老甚至死亡。为此,植物需要一个能够统筹调控抗病反应的基因决定什么时候该产生抗病反应,什么时候该关闭抗病反应。NPR1作为植物水杨酸途径的关键调控因子,不仅对植物系统获得抗性(systemic acquired resistance,SAR)和诱导系统抗性(induced systemic resistance, ISR)起核心调控作用,而且是植物基础抗性(basic resistance)以及抗病基因(resistance gene)发挥作用的决定性因素。NPR1作为多种信号途径的交叉点,既与TGA转录因子相互作用调控病程相关基因的表达,又与WRKY转录因子互作,在调节和平衡水杨酸和茉莉酸信号传导途径中起关键作用。
突变体信息
病程相关基因(PR)会在响应病原菌信号时强烈上调表达。顾名思义,npr1突变体在受病原菌侵染时,PR基因上调表达的现象完全消失,植物对病原菌的抗性降低,对水杨酸的响应减弱;在国内拟南芥突变体中心AraShare(http://www.arashare.cn/)可查询到2个株系,编号为N692710和N691675。

图1 npr1突变体对丁香假单胞菌的抗性减弱[1]
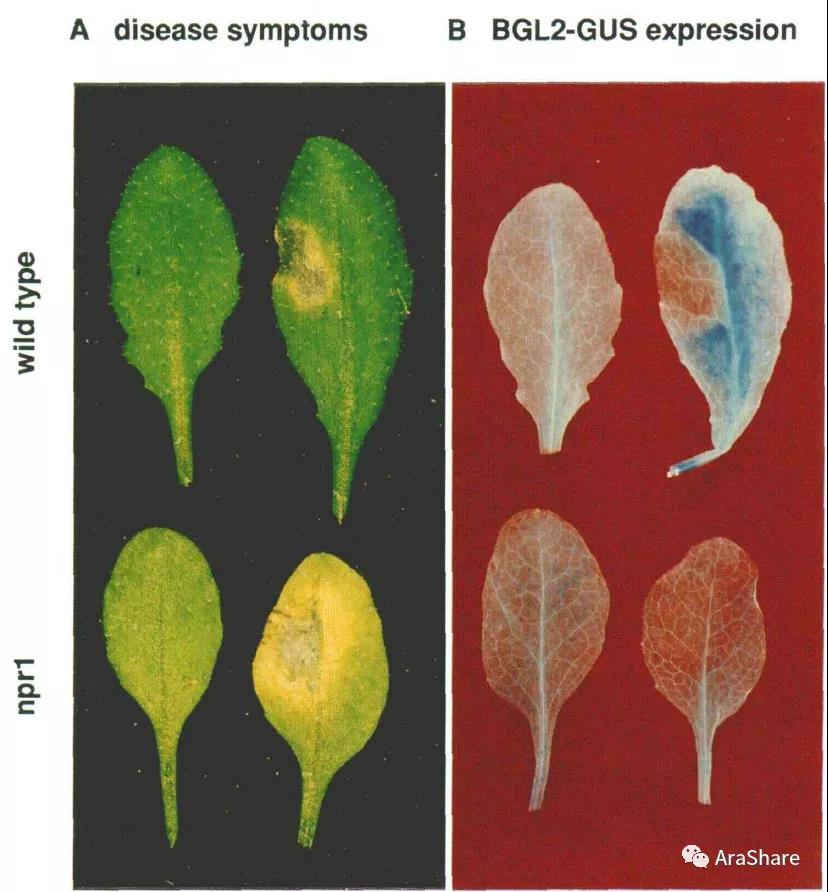
图2 响应病原菌时npr1突变体中抗性相关基因的表达减弱[1]
基因表达谱
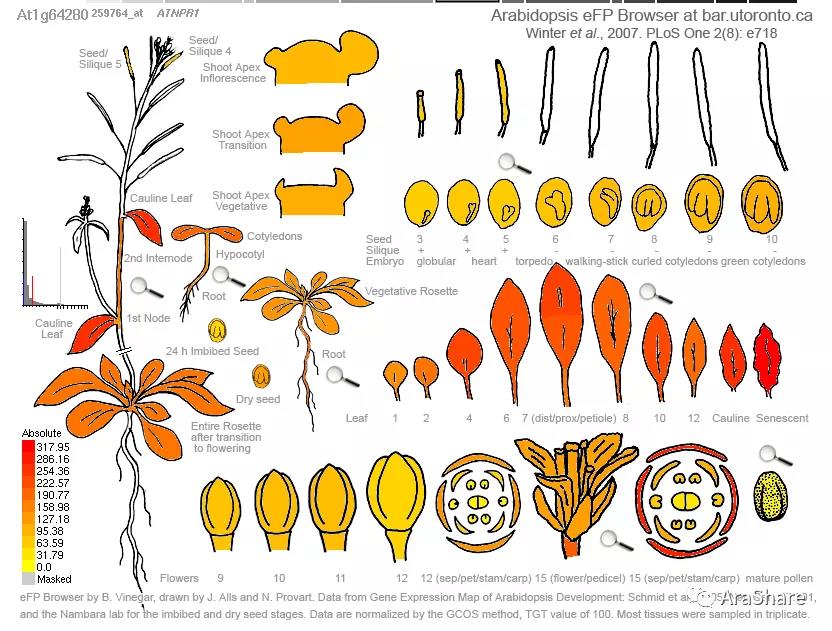
图3 拟南芥中NPR1 基因表达谱(来自eFP browser)
研究概况
1994年,现在的董欣年院士彼时还只有35岁。当时对植物免疫调节网络的研究刚处于萌芽状态,主要的研究方法是正向遗传学,也就是筛选突变体,通过筛选突变体来寻找植物抗病过程中的关键基因。但是,植物的抗性反应是看不见摸不着的,难道要对每一个要筛选的突变体都进行病原菌接种实验吗?这也太浪费时间和精力了!董院士精才绝艳,想出了一个解决之道。她在GUS前面连接了一个抗病相关基因的启动子,做成了转基因拟南芥。这个拟南芥在受病原菌侵染时会诱导GUS表达,最终叶片会表现出蓝色。对这个转基因植株进行诱变,再筛选叶片不变蓝的突变体,工作量就小得多了。通过这个巧妙的方法,董院士筛选到一个无法响应水杨酸的突变体,在水杨酸处理后PR基因的上调表达几乎完全消失。董院士给这个突变体起了个符合表型的名字:npr1(NONEXPRESSER OF PR GENES 1)[1]。三年后,经过董老师等人的不屑努力,终于通过图位克隆的方法克隆到了NPR1。NPR1编码一个含有ankyrin repeats结构域的蛋白,这个蛋白在动物中也存在同源基因,并且都参与抗病反应(动物中的NPR1同源基因参与aspirin途径)。于是,董老师揭示了一个动植物中保守的由NPR1调控的通路,该结果发表于1997年的Cell期刊上[2]。
在鉴定到NPR1以后,董老师发现NPR1不是转录因子,那么它又是如何调控PR1基因的表达的呢?为了回答这个问题,董老师利用NPR1作为诱饵,用酵母双杂交技术筛选到了拟南芥中NPR1的互作蛋白,发现了一个互作的bZIP转录因子AHBP, 它可以结合到PR基因的启动子上调控PR基因的表达[3]。
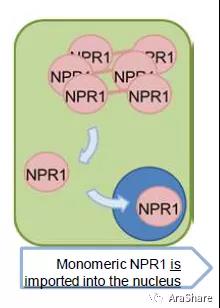
图4 NPR1由多聚体变成单聚体后入核的模型[4]
在发现了NPR1调控病程相关基因的机制是与转录因子互作以后,一个问题随之而来:NPR1如何被激活功能?又如何最终入核行使功能呢?原来,在没有病原菌信号的情况下,NPR1以多聚体的形式存在,而病原菌信号会导致植物细胞内的氧化还原力发生变化,最终使NPR1变成单聚体,从而可以通过核孔,最终调控抗病反应,该研究发表在2003年的Cell期刊上[4]。
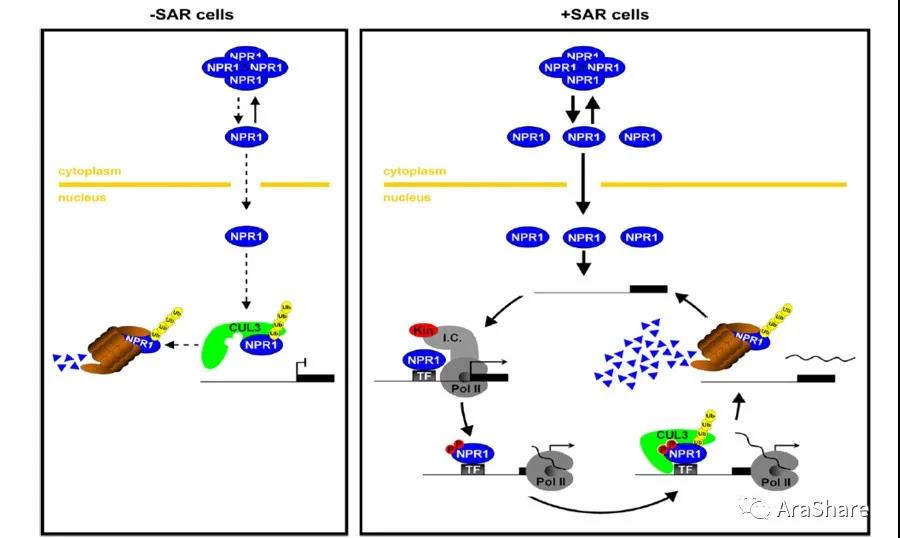
图5 NPR1被降解的模型[5]
九年过去了,至此,人们已经知道了植物如何踩抗病的“油门”,但是植物又如何“刹车”?要知道抗病相关蛋白的表达意味着生长所需的能量被急速消耗,如何刹车显得尤其重要。董老师带着这个问题又进行了探索,随着科学和技术的发展,蛋白的泛素化的检测变得越来越简便且经济,在2009年,董老师发现了原来NPR1在入核以后会很快的被26S蛋白酶体降解,最终导致抗病反应的停止[5]。
蛋白调控网络
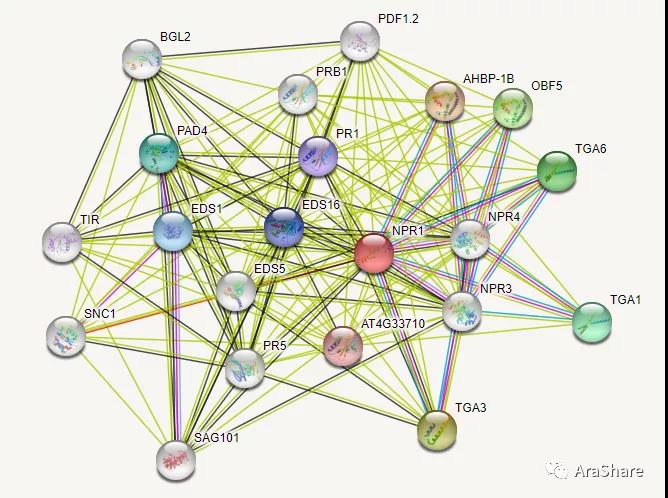

图6 拟南芥中NPR1蛋白互作网络预测(来自STRING)
如图6所示,NPR1互作蛋白网络已经得到比较好的剖析,NPR1与多个转录因子如TGA等存在互作,并且与PR基因,PDF1.2等典型的抗病相关基因存在共表达的关系。
其他物种研究情况简介
在真核生物中,NPR1同源蛋白的结构域具有较高的保守性。在动物中,含有ankyrin repeat结构域的蛋白也被报道可以调控基因表达,其中比较著名的是NF-kB的前体p105和抑制子I-kB。这两个基因参与动物的多个生理过程,包括抗病过程,同时受多种病原物的诱导,并且这种诱导可以被阿司匹林抑制。
展望
NPR1长期处于研究热点,截止至小编写完这篇稿子时,在2020年就已经有81篇NPR1的相关研究报道。这些研究不再局限于拟南芥,更拓展到各个重要的农作物,并且也不再局限于植物的抗病相关途径,而是发现了NPR1在更多生物学过程中的角色。最后,笔者列出近几年的几篇美文,以飨读者。
1.Opposite Roles of Salicylic Acid Receptors NPR1 and NPR3/NPR4 in Transcriptional Regulation of Plant Immunity. Cell. 2018
2.NPR1 mediates a novel regulatory pathway in cold acclimation by interacting with HSFA1 factors. Nature Plants. 2018
3.Salicylic acid-induced transcriptional reprogramming by the HAC-NPR1-TGA histone acetyltransferase complex in Arabidopsis. NAR. 2018
4.Dynamic ubiquitination determines transcriptional activity of the plant immune coactivator NPR1. PLNAS. 2019
5.Formation of NPR1 Condensates Promotes Cell Survival during the Plant Immune Response. Cell. 2020
参考文献:
[1] Characterization of an Arabidopsis Mutant That Is Nonresponsive to Inducers of Systemic Acquired Resistance. The Plant cell 6, 1583-1592 (1994).
[2] The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell 88, 57-63 (1997).
[3] Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. PNAS 96, 6523-6528 (1999).
[4] Inducers of Plant Systemic Acquired Resistance Regulate NPR1 Function through Redox Changes. Cell 7, 935-944 (2003).
[5] Proteasome-Mediated Turnover of the Transcription Coactivator NPR1 Plays Dual Roles in Regulating Plant Immunity. Cell, 2009 May 29;137(5)
本推文基于科学文献和网络资讯写作而成,未经领域内专家审阅;限于笔者学识水平,难免存在错漏,仅供参考!