
 敬请关注公众号
敬请关注公众号

基因信息
Locus:AT2G14610
Name:ATPR1
OtherName:ATCAPE9,PATHOGENESIS-RELATED GENE1, PR1, PR1
基因功能
病程相关蛋白PR是植物受病原物侵染或非生物因子刺激后产生的一类水溶性蛋白,参与了植物的诱导抗病性,主要功能是攻击病原物,降解细胞壁大分子,降解病原物毒素,抑制病毒外壳蛋白与植物受体分子结合等。拟南芥PR1的生物化学功能尚不清楚,目前研究表明其具有抗真菌和卵菌的活性。
突变体表型
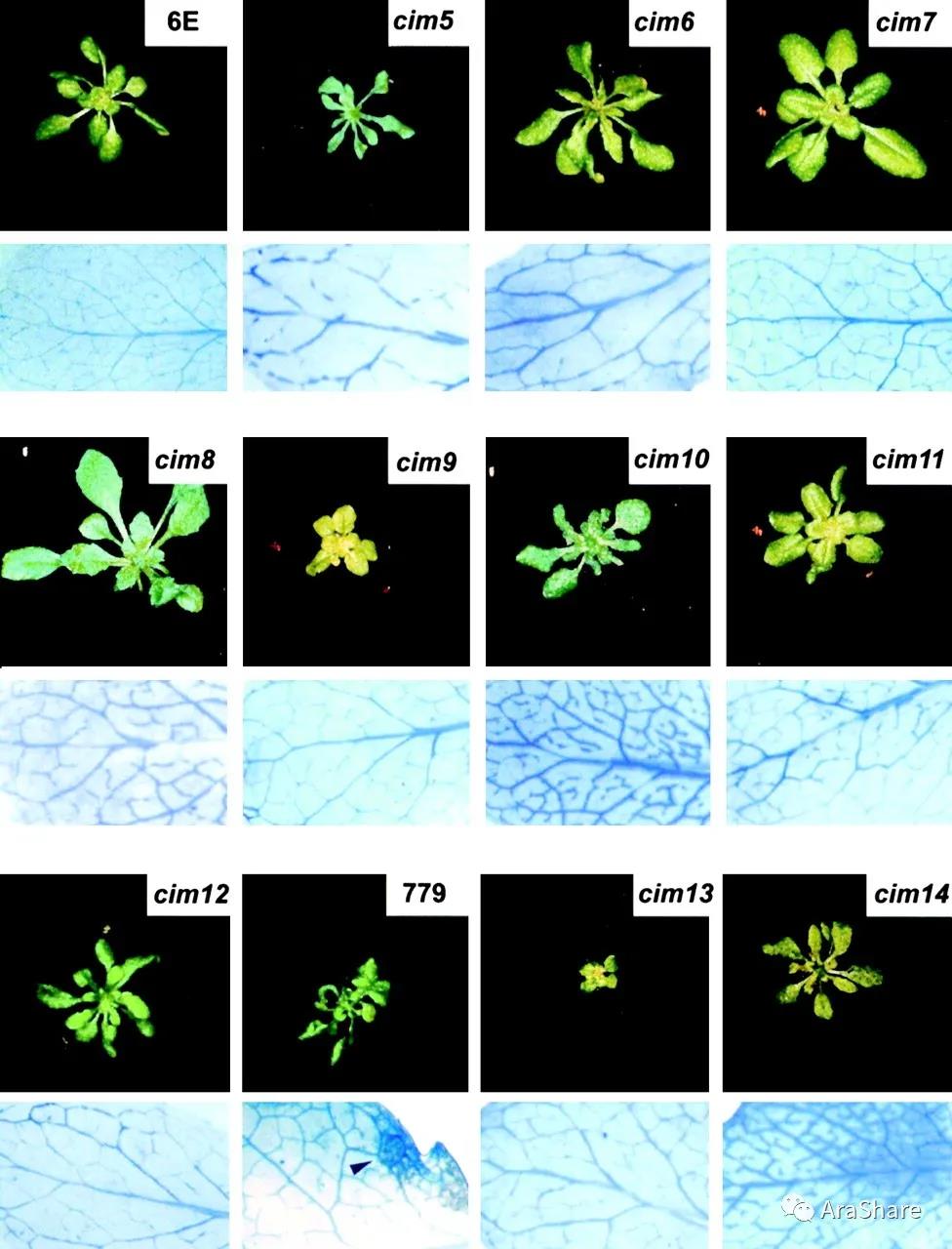
在TAIR官网上有PR1组成型表达突变体cim6的信息。该突变体信息来自著名植物学家Jeffery L Dangl的课题组2002年发表在Genetics上面的文章。为了鉴定组成型表达系统获得性抗性(SAR)的拟南芥突变体,他们构建了表达SAR诱导的PR-1启动子(PR-1/luc)控制的荧光素酶报告基因系。通过EMS诱变得到的抗病突变体不表现出自发的细胞死亡,但很多发生了形态改变,比如叶片窄小、卷曲,有的出现点状坏死斑。在AraShare突变体库中暂无该突变体信息,欢迎同行朋友捐赠。
基因表达谱
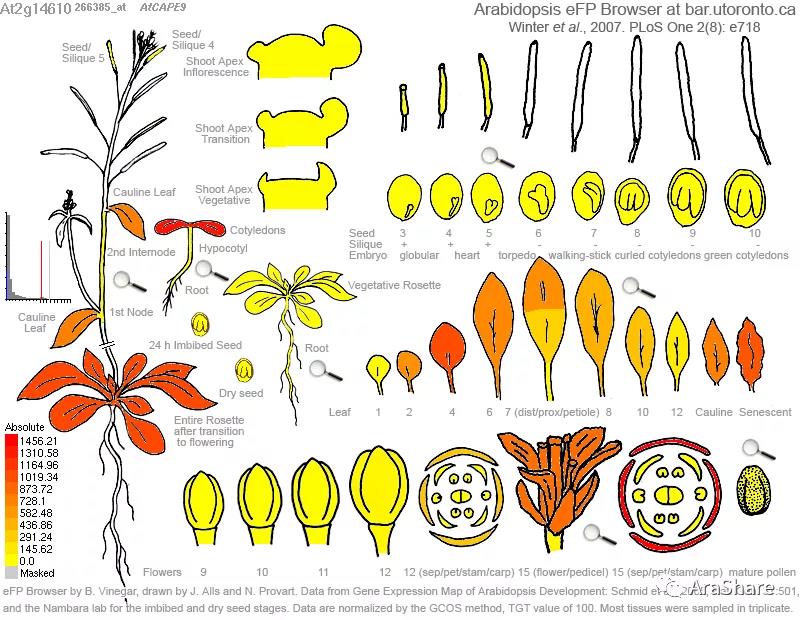
拟南芥中PR1基因表达谱(来自eFP browser)
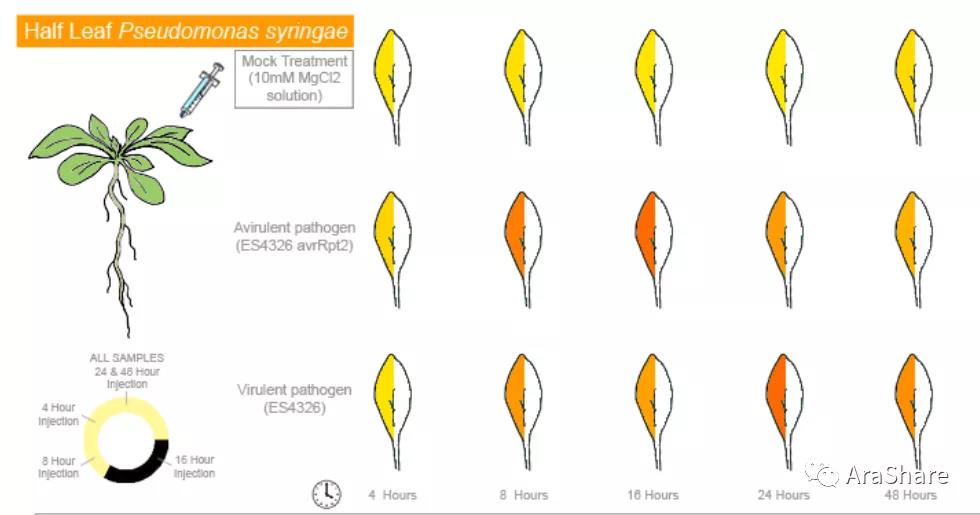
PR1受病虫害等生物胁迫诱导,如图所示,在丁香假单胞菌及其无毒株侵染后24~48小时内,PR1基因的表达量出现了明显的升高。
研究概况
1. 庞大的PR蛋白家族
1970年,美国科学家首次在感染TMV的烟草叶片中发现病程相关蛋白(PRs)。1979年,white等首次发现了外施的水杨酸或乙酰水杨酸能够诱导烟草对TMV产生全株抗性,并且也表达了TMV感染后生成的PRs[1]。之后的一系列研究证实了细菌、真菌、病毒侵染植物后产生的一些PRs,均可被水杨酸或乙酰水杨酸诱导。科学家发现抑制水杨酸合成的转基因植株,其系统获得性抗性(systematic acquired resistance,SAR)会受到抑制,从而将水杨酸认定为SAR反应的重要信号分子。顺理成章地,在免疫响应中普遍存在又反应灵敏的PR蛋白就成为了水杨酸介导的抗病反应的分子标记。
PR蛋白家族的划分在几十年间经历了不断的扩充完善,根据PR基因编码蛋白质的氨基酸序列相似性、血清学关系和酶分子的活性分为17个家族。
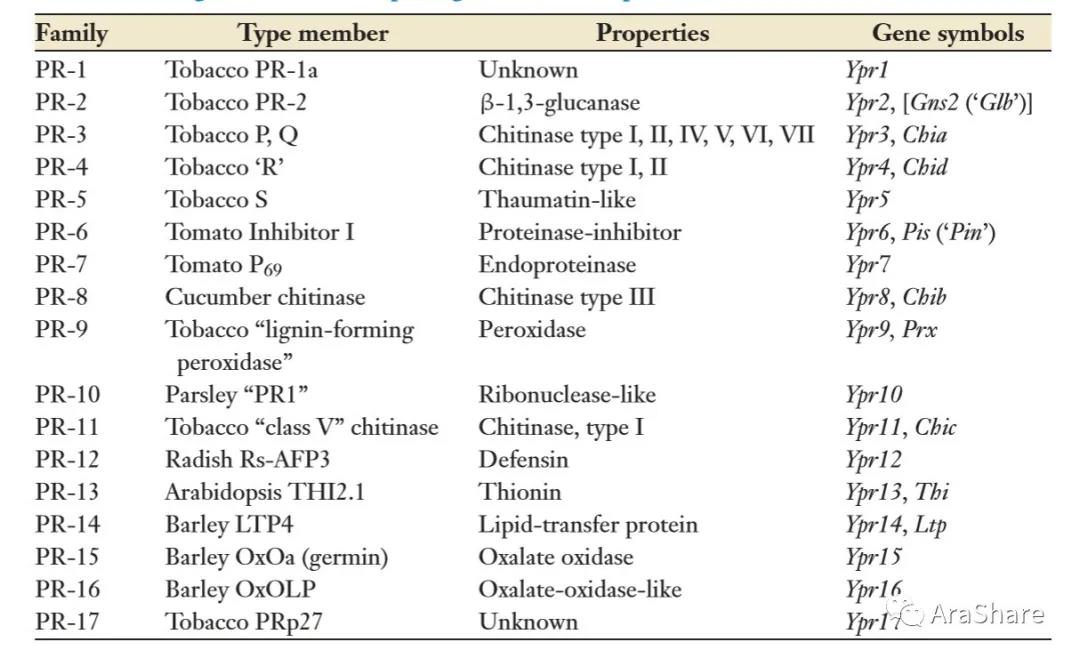
已知的病程相关蛋白家族[2]
PR-1家族在迄今所调查的每一种植物中都存在,所有的PR-1蛋白在结构上都是相似的,并共享一些严格保守的残基,包括6个半胱氨酸。这些发现表明,PR-1独特的分子结构在进化过程中高度保守,了解其抗病机理具有非常重要的意义。
2.“冒名顶替”事件
PR-1蛋白家族已经被广泛用作水杨酸(SA)介导的抗病反应的分子标记,但是拟南芥中PR-1基因的确定却经历了一场有趣的“顶替事件”。最初,拟南芥基因组计划将At2g19990注释为PR-1,该基因在两个拟南芥全基因组芯片中也被鉴定为PR-1, TAIR引用了大约60篇文章来描述其表达模式。然而,科学家们发现这些实验中使用的探针并不是At2g19990,而是一个同源基因At2g14610,被注释为PR-1-like。
为了让这位真正可靠的“分子标记”浮出水面,科学家们分析了这两个基因在拟南芥中的表达。At2g14610 (PR-1-like)表现出典型的水杨酸响应表达模式:在SA处理、丁香假单胞菌和CaMV侵染后,野生型拟南芥中mRNA水平升高。相反,At2g19990(注释为PR-1)的表达在SA处理的Col-0中未检测到,虽然它对病原体有反应,但并不显著。从而科学家提出应该为真正的PR-1正名,注释应该将At2g14610标识为真实的PR-1。At2g14610得以摘下“高仿”的帽子,而At2g19990也被打回原形。这项有意思的研究也被发表在当年的Planta杂志上。[3]
3. PR1的功能挖掘
根据L.C. van Loon等构建的拟南芥预测PR-1蛋白家族系统发育树,拟南芥基因组编码了具有预测PR-1结构域的22个蛋白。[2]在拟南芥中,PR-1家族同源基因大多在根和花粉中组成性表达,只有一个PR-1基因(At2g14610)被病原体、昆虫攻击或化学处理激活。
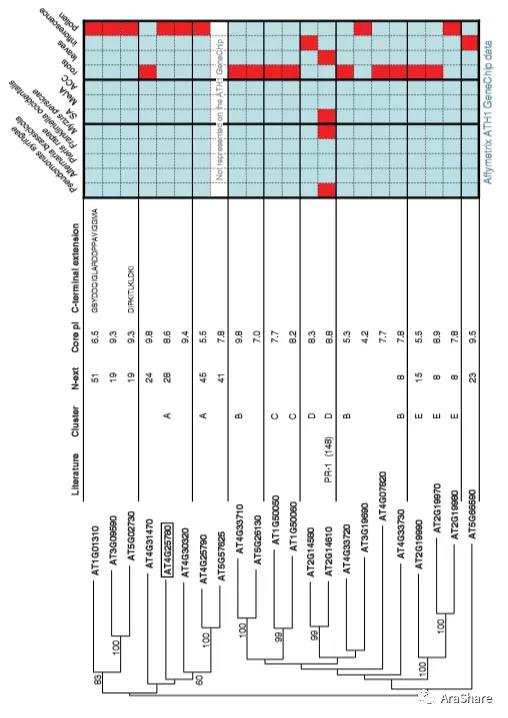
预测拟南芥PR1蛋白家族系统发育树
早在上世纪90年代,科学家们就发现拟南芥NPR1基因突变体表现出系统获得性抗性的受损,信号分子PR1的表达量显著降低。[4-5]种种迹象表明,NPR1或许就是调控PR1表达的关键因子。1999年,董欣年课题组发现NPR1与bZIP转录因子TGA相互作用。通常情况下,NPR1在胞质中是寡聚体的存在形式,单体入核后被磷酸化,与TGA家族的多个转录因子发生强烈互作,TGA转录因子结合到PR1基因的启动子区,诱导了PR1的表达,激活了SAR反应。[6]从此奠定了NPR1核心抗病基因的地位。
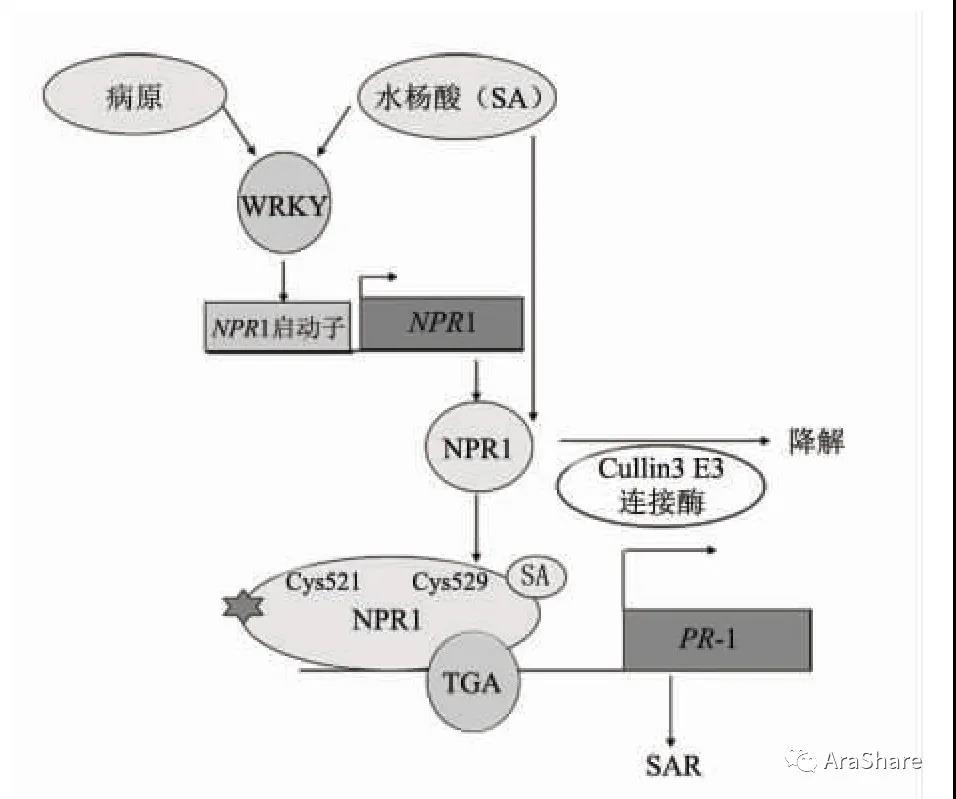
SA介导的NPR1抗病信号转导途径模式图
总体上,近年来对于引起PR1基因表达的信号传导及 PR1基因的表达调控机理的研究逐渐增多,尽管其分子机理还不够明晰,科学家已经利用转基因植物进行了很多PR基因相关的抗病育种研究,目前已卓有成效。
基因网络
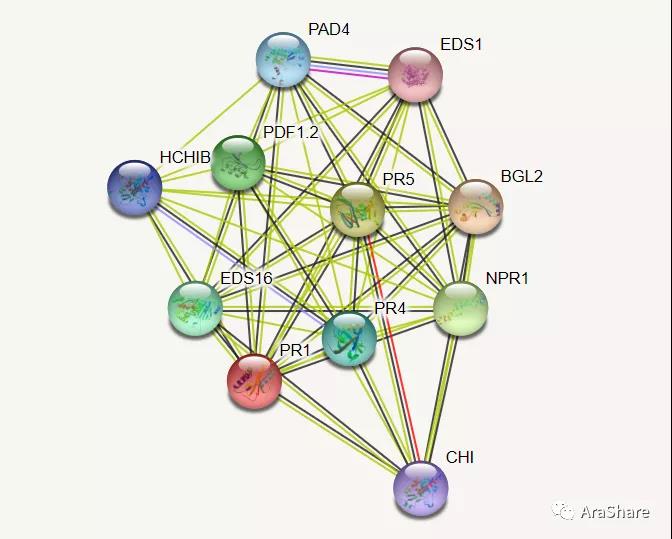
如图所示,STRING预测的与PR1有互作的蛋白网络,PR1与这些基因都存在共表达关系,其中EDS1/NPR1/PAD4/PDF1.2等都是经典抗病相关基因。
其他物种研究情况
简介近年来,尽管对于引起PR基因表达的信号传导及 PR基因的表达调控机理的研究并不非常明晰,科学家已经利用转基因植物进行了很多PR基因相关的抗病育种研究。除了烟草之外,水稻、小麦、番茄等作物中PR1蛋白的表达特性都得到了研究,人们可通过构建 PR蛋白转基因株系的方法获得广谱抗菌、作用持久的抗病新品种。例如Sarowar 等[7]对转化来自辣椒(Capsicum frutescens)的一个编码碱性PR-1蛋白基因的烟草植株进行了研究,结果表明这个基因在烟草中的表达,不仅增强了烟草对重金属胁迫的抗性,也增强了其对卵菌病原物烟草黑胫病菌(Phytophthora nicotianae)、细菌性病原物茄科雷尔氏菌(Ralstonia solanacearum)和烟草野火病菌(Pseudomonassyringaepv.Tabaci)的抗性。
展望
随着科学技术发展,对病程相关蛋白特性与功能的研究逐渐增多。国内对植物转化PRs的研究也十分活跃,特别是对农杆菌介导的几丁质酶和β-1,3-葡聚糖酶基因转化马铃薯、黄瓜和花生的方法进行了系统的研究。各类转基因植物的成功,使得某些植物在一定胁迫条件下的抗性有所提高。但对于特定分子机制的研究仍然有待深入,未来需要对病程相关蛋白的结构、功能以及参与的信号转导途径进行进一步的探究,阐明基因的表达调控规律和对靶标病原物的作用模式,解开PR蛋白的生物学功能之谜,从而有利于筛选出更加广谱高抗的植物群体。
参考文献:
[1] Acetylsalicylic acid (aspirin)induces resistance to tobacco mosaic virus in tobacco. Virology, 1979,99(2):410-412.
[2] Significance of inducible defense-related proteins in infectedplants. Annu.rev.phytopathol, 2006, 44(1):135-162.
[3] Inappropriate annotation of a key defence marker in Arabidopsis:will the real PR-1 please stand up? Planta, 2004, 219(6):1089-1092.
[4] Characterization of an Arabidopsismutant that is nonresponsive to inducers of systemic acquired resistance. The Plant Cell, 1994, 6 (11): 1583-1592.
[5] Isolation of Arabidopsis mutantswith enhanced disease susceptibility by direct screening. Genetics, 1996,143(2): 973-982.
[6] Interaction of NPR1 with basic leucine zipper protein transcriptionfactors that bind sequences required for salicylic acid induction of the PR-1gene. Proceedings of the National Academy of Sciences, 1999.
[7] Overexpression of a pepper basicpathogenesis-related protein 1 genein tobacco plants enhances resistance toheavy metal and pathogen stresses.Plant Cell Reports, 2005, 24(4):216-224.
本推文基于科学文献和网络资讯写作而成,未经领域内专家审阅;限于笔者学识水平,难免存在错漏,仅供参考!